 吉川泰弘の
ホームページへようこそ!
吉川泰弘の
ホームページへようこそ!
遠隔VODでやった授業をYou Tubeに載せる方法を学びました。1回から8回までの講義を全てZOOMで作成しました。結構労力がかかりました。せっかくなのでYou Tubeで紹介することにしました。興味のある方は見て下さい。全部で20分割されています。何回か講義の順番を変更したりしたので混乱が起きています。2021年の講義で整理しなおしました。
第1回は概要、地球環境の推移と生物、原核生物(細菌)の進化です。
概要(解説)https://www.youtube.com/watch?v=XPdjeHOviZI
地球環境 https://www.youtube.com/watch?v=pkd9nKv40Mc
真正細菌 https://www.youtube.com/watch?v=orwUICealGo&t=378s
古細菌 https://www.youtube.com/watch?v=79d2ynGN4_Q
第2回は最初の真核生物である単細胞の原生動物とそれに感染するウイルスです。
原生生物 https://www.youtube.com/watch?v=lsRPFVtpET0
共生 https://www.youtube.com/watch?v=2KyUyMsbXuk
原虫 https://www.youtube.com/watch?v=YUJw2tH-QQE
ウイルス https://www.youtube.com/watch?v=o6fAJUaiqdU&t=0s
第3回は単純多細胞生物群です、真菌から3胚葉の寄生虫(扁形、線形動物)までです。
真菌 https://www.youtube.co/watch?v=hOmmyBgWMxU
寄生虫 https://www.youtube.co/watch?v=RehWSBHlBow
第4回は無脊椎動物の代表として節足動物。海の甲殻類と陸の昆虫です。
概要 https://www.youtube.com/watch?v=9cTFEdMaLBY&t=0s
軟体動物 https://www.youtube.com/watch?v=oM5l0gfm5pY&t=0s
甲殻類 https://www.youtube.com/watch?v=hyrHHHiUWbc&t=0s
昆虫 https://www.youtube.com/watch?v=CM1ADBwP1Z8&t=0s
第5回は脊索動物(ホメオボックスの原点)、無顎類、魚類、両生類、爬虫類です
脊索動物 https://www.youtube.com/watch?v=p7Q-RU0Jw-w
魚類・両生類・爬虫類 https://www.youtube.com/watch?v=3-o8W52Fr7Q
ゲノムと進化 https://www.youtube.com/watch?v=D_IQa8R8k30
第6回は鳥類と哺乳類の比較、免疫系、胎盤構造について考えてみます
免疫・胎盤 https://www.youtube.com/watch?v=H7EbOvU5cjs
翼手目・視覚系 https://www.youtube.com/watch?v=4ZCctYVilhI
第7回は、霊長類に絞って、サル類、類人猿、ヒトへの進化です。ヒトの特性は?
霊長類・ゲノム https://www.youtube.com/watch?v=TwAY6mSSm3w
ネオテニー https://www.youtube.com/watch?v=m6LGuQGCvdA
脳と情報 https://www.youtube.com/watch?v=13EDnqrzzV4
第8回はまとめです。進化論から認識される世界とは?One World.
2021年秋①、久々に難しい問題の質問が来ました。以前にも何回か議論した、細胞膜のエーテル結合とエステル結合に関するものです。もう一度考えてみました。
第二回の予習をしていて疑問に思ったことがあったのでメールさせていただきました。細胞膜が古細菌だけエーテル型脂質であるとおっしゃっていましたが、エステル型脂質とエーテル型脂質では膜の性質はどのように変化するのですか。また、エステル型とエーテル型では細胞膜的にはどちらの方が優れているのですか。教えていただけたら幸いです。
もう一度まとめてみました。エーテル型脂質とエステル型脂質の違い・特性
① エーテル型脂質は、グリセロール骨格に炭化水素がエーテル結合した極性脂質。
② エーテル型脂質を極性脂質として有する生物は古細菌と一部の好熱性細菌のみ。
③ 他の生物(真正細菌、真核生物)の有する極性脂質は2分子の脂肪酸がグリセロールに
エステル結合した構造。
想像をたくましくすると、地球が冷える前に繁栄した化学合成独立栄養の嫌気性・好熱菌は、エーテル結合型脂質膜(炭化水素+グリセロール)を有していた。38億年前に分岐した古細菌はその末裔。その後、地球が冷えてきて、炭酸ガス、酸素が使いやすくなると真正細菌にエステル型の脂質膜(脂肪酸+グリセロール)を持つものが出現した。酸素がより多くなり、冷えていく地球環境では、エーテル結合よりも、より反応性の高いエステル結合の脂質の方が適していたので、その後に出現した真核生物群はエステル結合型がメジャーになった?地球の温度が低下し、大気、海に酸素が増加したことと関係している?
炭化水素→エーテル結合→エステル結合:C-C-C-C→C-C-O-C-C→C-CO-O-C-C
インターネットで探してみたら、https://www.jstage.jst.go.jp/article/kagakutoseibutsu1962/28/5/28_5_288/_pdf/-char/jaに、詳細なエーテル型脂質に関するレビューがありました。僕の考えたほど単純ではないようです。
考察;「細菌がエーテル脂質を持つということは,古細菌にとってはどのような意義があるのだろうか.古細菌には,前述のように高度好塩菌,硫黄依存好熱古細菌およびメタン生成細菌(ユーリー古細菌群)があるが,これらの菌は互いに生育環境も,特有のエネルギー獲得様式も異なっているので,共通した意義を見つけだすのは困難である.化学的に安定な飽和アルキル基のエーテル結合は,太陽光のもとで空気(酸素)とともに生育する高度好塩菌や,高温と酸性条件に耐えなければならない好熱好酸菌にはより適切なものかもしれないが,多くのメタン生成細菌にとっては さほど必要とは思われない.グリセロールのsn-1位に極性基が結合した構造は,他の菌が生産するホスホリパーゼ類に対する対抗策と考えられないでもないが,多くの真正細菌と共生しているメタン生成細菌にとっては有効でも,通常の菌の棲息できない異常環境にある高度好塩菌やイオウ依存好熱菌には必要とは思われない.テトラエーテル型脂質は,以前は高温環境下に適応するためと考えられていたが,好熱性でないメタン生成細菌であるMethano-brevibacter arboliphiticusにも,好熱性のMetha-nobacterium thermoautotrophioumと同程度の量のテトラエーテル型脂質があり,イオウ依存好熱古細菌であるThermococcusはジエーテル型脂質のみからなることを考慮すると,高温環境に適応するためとは考えにくい.このように,現在の古細菌の生育環境条件とエ ーテル脂質の存在を統一的に理解することは困難である.そこで,エ ーテル脂質の存在意義について2つの考え方ができる.①一つは,古細菌の祖先が発生した時点では何らかの理由によりエーテル脂質の採用は一つの有効な選択であったという考え方である.②もう一つの考え方としては,エーテル脂質もエステル脂質もともに膜形成能力を持っており,細胞発生初期にエーテル脂質を使った細胞(古細菌)とエステル脂質を使った細胞(真正細菌)が並列して発生したが,エーテル脂質を持った菌は他の生物の侵入しにくいところにだけ生き残った,というものである.後者はエーテル脂質を適応の結果とみるよりも,むしろ進化の遺物とみる見方である.この問題の解決のためには,エーテル脂質の生化学,生物物理学,古細菌の系統と進化など,広い範囲の研究結果に基づく総合的考察が必要であろう。」ということでした。
2021年春①VODの授業から始めましたが、学生さんから質問が来ました。これまでにない面白い質問を挙げておきます。

真正細菌が住み良い環境に生息して、メタン菌や好熱菌、好塩菌などの古細菌は極限環境に生息しているものが多いとありました。どういう点で、真正細菌のほうが住みよい環境での生存競争に有利だったのか教えてください。
長い細菌界の歴史(40億年)の中で地球環境は随分と変化しました。環境温度や海水塩分などが下がったこと。②シアノバクテリアの登場以来、酸素濃度が上がったこと。③真核生物が出現したこと。④高等多細胞動植物が出現し、地球のメジャーな生物群になったことなどが影響しています。こうした環境変化に適応したのが真正細菌群です。適応性が低かったのが古細菌でしょう。アシナシイモリ、ムカシトカゲ、シーラカンス、オウムガイ・・・といった生きた化石のような細菌群ということになると思います。従って、古細菌群には、①好熱菌や高塩菌が多い、②光合成能を持つ古細菌はいない。③適応した古細菌は、多分、真核生物の母体になった?発展した古細菌は原核細胞から真核細胞に化けた?④後の環境で出現した、後発の動植物と共生する機会が少ない、といったことだと思います。
2021年3月、全部の授業が終わってから2020年春①の学生さんから久々の質問がありました。面白い質問でした。載せておきます。○○○○です。
質問内容:グラム陽性菌である黄色ブドウ球菌(コアグラーゼあり)にあるコアグラーゼは血漿凝固作用を有していることを調べました。この時にコアクラーゼの反応で黄色ブドウ球菌は凝固した血漿で自身を包み免疫反応を回避するということもわかりました。その後に吉川教授のホームページで耐性菌AMRのところの載っているホスホマイシンが黄色ブドウ球菌の抗菌剤として用いられていることが分かったのですが、血漿に包まれている黄色ブドウ球菌にホスホマイシンが抗菌剤として使えるのでしょうか?
A:黄色ブドウ球菌
フィルミクテス門, バシラス綱,バシラス目,ブドウ球菌科,ブドウ球菌属の黄色ブドウ球菌。通性嫌気性のグラム陽性球菌。顕微鏡で観察すると、ブドウの房のように複数の細菌が集団を形成しているのでこの名がある。他の細菌と比較して高濃度 (10%) の食塩存在下でも増殖が可能であり、カタラーゼ活性、ブドウ糖発酵性を持つ。
B:黄色ブドウ球菌感染症
軽度のものから生命を脅かすものまで幅がある。最も一般的なブドウ球菌感染症は、①皮膚感染症(しばしば膿瘍を引き起こす)②しかし、細菌が血流に入り(菌血症と呼ばれる病態、敗血症)、体内のあらゆる部位に広がることもある。③その場合、心臓弁(心内膜炎)と骨(骨髄炎)によく感染する。
C: 黄色ブドウ球菌の病原因子
細胞壁に局在する病原因子: ①プロテインA - 細胞壁に存在するタンパク質で、黄色ブドウ球菌のほとんどが有する特徴的な成分。抗体のFc領域に結合する性質を持ち、これによって抗体の持つ生物活性を抑制することで、菌が免疫系によって排除されることを防ぐ働きを持つ。②フィブロネクチン結合因子 - 細胞壁に存在するタンパク質で、フィブロネクチンと結合して体内に定着する働きを持つ(定着因子)③タイコ酸 - 細胞壁に存在する分子で宿主細胞との結合を高める(定着因子)。
外毒素:① エンテロトキシン群 - 食中毒の原因となる黄色ブドウ球菌が産生する。下痢や腹痛などの直接の原因になるほか、嘔吐中枢にも作用して嘔吐の原因にもなる。スーパー抗原としての活性を持つ。②TSST-1(毒素性ショック症候群毒素-1) - 毒素性ショック症候群の原因となる毒素。強いスーパー抗原活性を持ち、発熱や悪心、ショック症状を引き起こす。免疫系をかく乱する役割を果たす。③表皮剥脱毒素 - スーパー抗原の一種。④溶血素- 赤血球を破壊する溶血活性を持つ毒素群、特にα毒素が重要な病原因子。免疫細胞を破壊することで菌の排除を防ぐ働きを持つ。組織破壊によって病巣部から周辺組織に侵入する際にも働く。⑤ロイコシジン - 白血球を殺す毒素であり、免疫細胞の破壊によって菌の排除に対抗している。
酵素群:①コアグラーゼ、クランピング因子 - 血漿を凝固させ、フィブリン形成を起こす。これによって菌の増殖の場となる凝集塊を作り出し、白血球や血漿中の抗体による排除を防ぐ働きがあると考えられている。②スタフィロキナーゼ - 析出したフィブリンを溶解させる働きを持つ。菌が凝集塊の中で増殖した後、その凝集塊を分解して周囲に感染を広げる際に働くと考えられている。③プロテアーゼ(蛋白分解)、④DNase(DNA分解)、⑤リパーゼ(脂質分解) -周辺組織を分解して感染の拡大に関わる。
D: ホスホマイシン
ホスホマイシンは、ストレプトミセス属の真正細菌が産生する抗菌スペクトルの広い抗生物質の一つで構造は極めて単純。作用機序は、β-ラクタム系抗生物質が細胞壁のアラニンのペプチド鎖を標的とする最終段階での阻害薬であるのに対し、ホスホマイシンは極めてユニークで、細菌の細胞膜能動輸送系によって効率的に菌体内に取込まれ、細胞壁peptidoglycan(ムラミン酸とアセチルグルコサミンの糖鎖)の生合成を初期段階で阻害することにより抗菌作用を示す。
解答
ブドウ球菌の生体内増殖は多様です。確かにコアグラーゼはフィブリン形成を足場に菌が生体防御系から逃げるように働くようですが、本来、異物が入ったときは生体防御系はフィブリンなどで包もうとします。菌が包まれたまま持続しようとすれば、いずれ好中球やマクロファージ、リンパ球が来て戦いになります。細菌がジッとしていないで積極的に増殖しようとするときは、上述のように自分の酵素でフィブリンを溶かして周囲に広がらなければなりません。危険覚悟で拡散するわけです。ホスホマイシンは細胞壁の厚いグラム陽性菌には、特に有効です。抗生物質の作用機序については、You Tubeを見てください。
https://www.youtube.com/watch?v=XO7JQ3otnjY
期末試験が終わりました。初めてのVOD講義でした。それなりに盛り上がっていたので期待しましたが、結果的には例年通りの成績でした。考えてみれば、いつも対面授業でやっていることをVODに変えただけなので、理解力が急に上がるわけはなかったと反省しています。VODと対面授業を組合わせて両方の長所が生かせるような授業をしてみたいと思っています。面白かった質問6をアップします。今期春①はこれで終了です。春②には動物感染症(獣医3年)、VPPの獣医関連法規(1年)、人獣共通感染症(VPP2年生)、獣医関連法規(3年)と千葉科学大学の人獣共通感染症(zoom)と授業の目白押しです。
面白い質問がありました。今回獣医学科の3年次生の「動物感染症学」で軟体動物の感染症を説明した時に尋ねられた質問です。こちらの方があっているのでこちらに載せます。
タコの感覚器系のところで、タコには涙孔があると記載されていました。しかし、タコは海に生息しているのにどうして涙孔が存在するのでしょうか? 常に水にさらされているのなら涙を放出する必要がないように思えました。面白い質問有難う。試験問題作成をしなければならないのに、つい気になって調べてみました。
1.タコの眼の前方には涙孔(lacrymal pore)が開孔する。
Lachrymal, lacrymal, lacrimalは同義で涙の意味
2、イカの眼は頭部両側に位置し、角膜、虹彩と虹彩に囲まれた瞳孔、水晶体、網膜、及び、視神経から成る。開眼類では、角膜は丸く開いており、眼窩の前部は海水が自由に流入する。他方、タコのような閉眼類やコウイカ類では、角膜は殆ど閉じており、涙孔と呼ばれる小孔が眼窩前部と通じている。
3、とすれば、開眼類では、海水が前眼房水と直接混ざることになります。他方閉眼類では、我々と同様に角膜は閉じている?ので、涙腺は涙孔として瞼と角膜の間に開口する。従って涙孔から出る涙(体液)は海水と混ざることになります。海水の塩濃度は、約3.2%、われわれのように体液が生理食塩水と同じなら0.9%、浸透圧順応型なら3.0~3.4%位でしょう。①前者なら浸透圧差(体液と海水)を少なくして角膜を保護する。あるいは②水圧変動(水深)と眼圧の差の調節に涙腺を利用する?そんなことが出来るか分かりませんが。③一部の魚類と同様に涙腺に唾液腺と同様に餌をとるための麻痺性毒液を入れている、などという説もあるようです。
4、また、涙孔と呼ばれる小孔は眼窩前部と通じ涙腺に繋がっているはずですから、海水と体液のやり取りができるように小孔となって開閉しているのではないでしょうか?そうだとすると、涙腺で何か環境変化を監視する仕組みがあるかもしれません。しかし、明確な説明は英文でも調べましたがありませんでした。
5、上記の浸透圧差の考え方は間違いでした。頭足類は浸透圧順応型で、体液の浸透圧は海水と同じのようです。「一般的なタコを含む頭足類の血リンパ、心外液および尿は、すべて相互に、また周囲の海水と等浸透圧です。頭足類は浸透圧調節しないことが示唆されており、これは彼らが浸透圧順応型であることを示します。これは、環境の浸透圧に合わせて適応することを意味します。浸透圧勾配がないため、生物から海水への、または海水から生物への水の純移動はありません。タコの平均最低塩分要件は27g/l(2.7%)であり、環境にかなりの量の淡水をもたらすいかなる障害も致命的となる可能性があります。」ということでした。従って仮説①はありません。
なお、地中海のタコの唾液腺から抽出したeledoisin(11アミノ酸、undecapeptide)が、涙の分泌を促進するという研究が報告されています。構造式はpGlu-Pro-Ser-Lys-Asp-Ala-Phe-Ile-Gly-Leu-Met-NH2です。また、涙管の進化については、以下の記載がありました。また
The earliest creature with a nasolacrimal duct is thought to have been Eusthenopteron or Osteolepis both of which were considered ancient lobe-finned fish.44 The lobe-finned fish include extant lungfish and the relict coelacanth. 鼻涙管が最初にみられるのはユーステノプテロン、オステオレピスでどちらも条鰭類で肺魚やシーラカンス(肉鰭類)を含むグループです。もっとも、軟体動物は魚類とは別に進化しているので、どこまで共通かは分かりません。
おまけ:軟骨魚類が浸透圧順応型で尿素を体内にためているのは知っていましたが、軟体動物の多くが浸透圧順応型であることは知りませんでした。彼らがどのように高浸透圧適応をしているのかは、試験が終わったら調べてみます。ありがとう、面白かったです。













今回、8回目の講義スライドに加えて新規に「環境と生物進化」を追加しました。何度もの大量絶滅の危機を乗り越えてきた生物の末裔が、現在の世界を作っていることを理解してもらおうと思ったからです。また、生物の多様性を維持することが何故必要か?を理解してもらおうとも思ったからです。このような歴史と世界を理解できるようになった人間とは何者なのか?また、どうすべきなのか?を考えてもらいたかったからです。








2020年4月20日から「新型コロナウイルス感染症COVID-19」の流行拡大により、この授業もVOD(Video On demand)方式に変えました。取りあえず順調?に進んでいます。学生さんは、いつも通り、面白い or 考えさせられる質問を送ってくれます。講義への面白い質問と回答です。連休中に名言をみつけました。
少而學、則壯而有爲 少にして学べば、則ち壮にして為す有り
壯而學、則老而不衰 壮にして学べば、則ち老いて衰えず
老而學、則死而不朽 老にして学べば、則ち死して朽ちず すごい言葉です。
講義への面白い、考えさせられる追加質問への回答です。こちらを先にあげておきます。


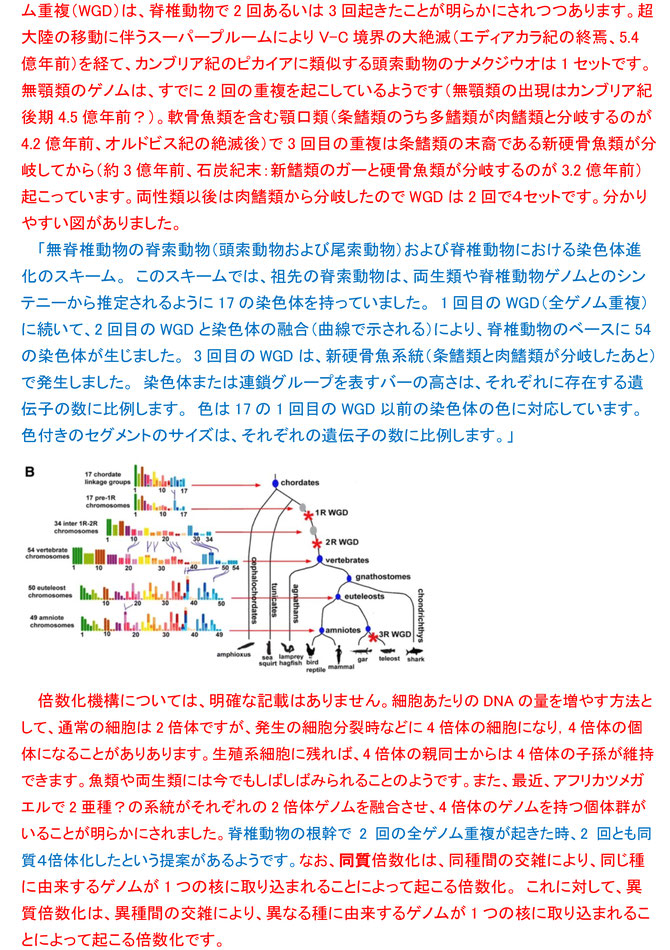







面白かった質問の4です










面白かった質問その3です。







第3回の講義(本日のVOD講義)に関する質問です。勉強になりました。






面白かった質問のまとめ1です。第1回と2回の講義についての質問です。






現代生物進化の2019春①が終わり, 早くも秋①が始まりました。2018年のスライドと質疑(現代生物進化の項参照)を見たうえで、講義を聞き、理解できたこと、理解できなかったことを、一つずつメールで送ってもらい、回答することにしました。2018年度は語句の質問と説明が多かったのですが、今回は、質問内容が深くなって回答に苦しむことが多くなりました。
また、説明とともに新しいスライドを追加することになりましたが、このホームページの「現代生物進化」の項目の容量が一杯になったようで、新しいスライド等を入れる余地がなくなったようです。仕方がないので、ここに「現代生物進化2」を開きました。時間を見て、7回目までの質疑をアップしいと思います。なお、2019年春①の試験問題と模範解答は一番後ろに載せてあります。
秋①の講義が終わりました。試験の回答を載せておきます。確認してください。
正しい文章には〇を、間違っている文章は、下線部を正しい言葉に置き換えなさい。
正しい文章は1つだけです。2つ以上〇をつけると減点になります。
第1回講義から
1、従属栄養細菌は他の生物が作った無機物を栄養源とする細菌である。( 有機物 )
2、真正細菌と古細菌の細胞膜は脂肪酸とグリセロールがエステル結合している。(真核生物)
3、ホイタッカーの5界説でモネラ界に属するのは原核生物である。( 〇 )
4、アルコール発酵が細菌によるものであることを明らかにしたのはロベルト・コッホである。(ルイ・パスツール )
5、古細菌のハロバクテリア綱には、その名の通り好気性の好熱菌が含まれる。( 好塩菌 )
第2回講義から
1、単細胞の真核生物である原生生物のうち葉緑体を持ったのは原生動物である。( 藻類 )
面白い回答、ミドリムシ(あるいはハテナ)間違いではありません。
2、1967年、細胞内共生説を唱えたのはマーギュリスである。( 〇 )
3、大腸バランチジウムは胞子虫門に属する。( 繊毛虫門 )
4、トリパノソーマは2宿主性である。鳥類・哺乳類を宿主とする種のもう一つの宿主はヒルである。
(吸血節足動物)
5、マラリア原虫で母細胞の分裂により多細胞となった状態の細胞塊をメロゾイドという(シゾント)
第3回講義から
1、担子菌門の約半分の種は、藻類との共生体を形成して地衣類として生活している(子嚢菌門)
2、動物の遺体は主として細菌が分解し植物の遺体は主として原生動物が分解する傾向がある。(真菌類)
3、ヒドラは、多細胞の動物の中で最も原始的な1胚葉性動物であり、器官を持たず左右の区別も
ない。(カイメン )
4、回虫など線形動物の多くは、雌雄異体であり、複数回の脱皮で成長する。( 〇 )
5、吸虫や条虫の多くは、中間宿主内で有性生殖をおこなう。( 無性生殖 )
第4回講義から
1、カンブリア紀の最大動物で最高の捕食者で節足動物といわれているのがアノマロカリスである
(〇)
2、旧口動物(前口動物)は、原口が肛門になって発生する動物群である。( 口 )
3、エビでは心臓は背側に、神経系は神経鎖となって腹側に、消化器系は体の背側に分布している。( 中央 )
4、エビの血液細胞には赤血球はなく,セミ顆粒球,ヒヤリノ細胞とリンパ球の3種類がある(顆粒球)
5、雄バチから放出されるフェロモンには階級社会を維持する物質がある。9-オキソデセン酸が有名である。(女王バチ)
第5回講義から
1、ナメクジウオは尾索動物であり、1から13までのホメオポックス遺伝子をすべて揃えている。(頭索動物)
2、ヌタウナギは無顎類に属し、嗅球、大脳半球、間脳、延髄などを持つが中脳を欠いている。( 小脳 )
3、ホヤは幼生時、オタマジャクシのような形をしているが、成体は岩などに固着しセルロースを合成する。( 〇 )
4、大部分の軟骨魚類は体液や細胞内に尿酸を蓄積し、体内と海水との浸透圧差を小さくしている。( 尿素 )
5、肺や気嚢の原点となった鰾(ウキブクロ)は魚類時代に鰓の一部から生じた。( 食道 )
第6回講義から
1、魚類ではゲノムを倍加していった。ヒトのゲノムの30倍以上のゲノムサイズを持つ魚はギンザケである。(肺魚)
2、異なる種の染色体間で、遺伝子が同じ順番で配置されていること、もしくはその領域をネオテ
ニーという。(シンテニー)
3、恒温動物とは外温性動物のうち、自律的に体温を制御している動物をいう。(内温性)
4、胎盤を介して抗体(IgG)が胎子に移行できない動物の代表は偶蹄類である。( 〇 )
反芻動物も正解です
5、ココウモリは超音波をだしエコロケーションをする。視覚系では立体視が出来る(立体視できない)
第7回講義から
1、約600万年前にチンパンジーから分岐した人類の祖先は猿人、原人、旧人、新人を経てホモ・サ
ピエンスとなった( 〇 )
2、ヒトの染色体数はチンパンジーより1本少ない。チンパンジーの第12染色体と13染色体が融合
し、ヒトの第1染色体となったためである。( 第2染色体 )
3、 アジア人、ヨーロッパ人のゲノムの1~1.5%はネアンデルタール人由来であるが、、アメリカ人の場合は0.1%以下。( アフリカ人 )。
4、齧歯類は未熟児として生まれる巣籠り動物であるが、ヒトは社会的に未熟な第2次巣立ち動物である。( 第2次巣籠り動物 )
5、ヒトで特に肥大した大脳皮質の部分は頭頂葉の前野で、自意識、意志、推測、やる気、戦略などを司っている。( 前頭葉 )
8, 面白かった質問から
1、核膜は内膜と外膜をもつ脂質2重層の4重膜構造をとっており外膜は小胞体につながっている。
(2重膜構造)
2、 マラリア原虫は無性生殖するが、ガメートゴニ―を行い有性生殖もする。マラリア原虫の終宿
主はハマダラカである。( 〇 )
3、ヒト胎児は尿を産生し、排出された尿は卵黄となる。卵黄には肺の成熟を促す物質が含まれており、胎児は卵黄を飲むことで、肺胞の発達を促している。( 羊水 )
4、哺乳類のゲノムは、祖先のナメクジウオのゲノムを1倍体とすると8倍体となっていると考えられる。( 4倍体 )
5、 最近、ジャンクDNAと考えられていたエキソンがmRNAを核から運び出す過程に関与していることが報告された。( イントロン )
講義を聞いた感想、あるいは予習・復習をどの程度やったか?質問への回答など、授業に関する自由な意見を書いてください。今後の授業に生かしたいと思います。
現代生物進化第1回質問回答まとめ(質問中、主なものを掲載しました)。
第1回講義真菌は体外消化を行い、植物よりは動物よりに分類されるとおっしゃいました。なぜ、見た目は植物に近くなったのでしょうか。面白い質問です。それは、真菌の生活様式が植物的だからです。①動物と植物の決定的な違いは、生命を維持するために動く生き物か(animal)、動かない生き物(phyto)かです。単細胞真核生物(原生生物)の時に光合成する(独立栄養の)藻類と従属栄養の原生動物類に分かれました。多細胞化した時に植物系と動物系に分かれましたが、真菌は動かないで従属栄養生活をする生物です。②我々の周りでも、家、ビル、教会、寺院のような建築物は動きませんし、そうした構造です。バイク、自動車、電車、飛行機などは動くもので、そうした共通構造を持ちます。生物も同じです。建築物は作ってしまえば、それだけですが、動くものは常にガソリンや電気のようなエネルギーが必要です。③こうしたたとえをすると、真菌類は、絶えず維持にエネルギーのいる建物ということになります。火力発電所や原子力発電所みたいなものでしょうか?④ちなみに、我々の直接の祖先になる、後索動物のホヤは、幼生期には魚のように泳ぎます。しかし、大人になると植物のように岩にくっついて生活します。植物からセルロース合成酵素をもらってセルロース合成をする唯一の動物です。面白いですね。

【理解できなかったこと】理解できなかったこととはまた少し違うような気もするのですが、吉川先生に聞きたいことがあります。高校生の頃に生物の授業の中で進化について勉強していました。その中でカンブリア紀の大爆発の代表的な生物としてアノマロカリスについて学びました。アノマロカリスは当時最大で最強の生物であると言われていますが、当時調べた本の中ではアノマロカリスは三葉虫を食べることができない(硬くて)ほどの弱者だったということが書いてありました。かつ触手のような頭から生えている2本の手は実際に曲げると口までは届かないということも聞きました。ではどうやってアノマロカリスは当時最強な生物になれたのでしょうか?進化という言葉を聞くといつもこのことばかり考えてしまいます。今日の4回目の講義で少しは答えられたと思います。古生物を含めて、その生き物の①実物の大きさと運動の能力、②生存時期・生存期間、③周囲の環境、④地球規模での環境変化を知らなければなりません。エディアカラの最後からカンブリア紀の初期にかけて、大きさ、運動能からみて、やはりアノマロカリスは最強の捕食者であったと思います。ただカンブリア紀は、全ての動物種が一気に登場したので、アノマロカリス(節足動物の甲殻類の祖先に近い?)とほぼ同時期に、強力な軟体動物のオウムガイの祖先や三葉虫が出てきます、また脊椎動物の進化も早く、シルル紀までには魚類の前の鎧を着たような板皮類が出現します。アノマロカリスが長く繁栄できなかったのは、同じ空間を生きる強力なライバルが出現し、①食物連鎖のトップを維持できなかったこと、②深海や淡水に逃げてすみ分ける能力がなかったからでしょう。もう少し早く出現していれば、長く繁栄したかもしれません。その意味ではカンブリア紀は、極端に弱肉強食の食物連鎖が進んだ時代だと思います。そのため多様性が広がり、進化が早まったように見えるのではないでしょうか(生物爆発)。それは、オゾン層や酸素濃度、好気呼吸のエネルギー効率、氷河期のあとの地球の安定的な温暖化などと関連しているかと思います。

メタン菌や好熱性菌などの古細菌は、分岐した時期の地球の環境が厳しく極限環境に生息しているようですが、長い年月を経た今の時代にも生息しやすい場所に移動しないで、わざわざ極限環境にいるのはどうしてですか。また、古細菌はその場所でどのような役割を担っているのですか。① 極限状況の環境では敵対する微生物が、ほとんどいないというメリットがあります。②また、一般的に生息しやすい?環境は、すでに生存競争に勝ち抜き進化した生物群が占めていて、生物学的ニッチ(隙間)がありません。③既に、極限環境に適応しているので、彼らにとって住みにくいことはないと思います。役割というよりも、その環境で栄養、代謝に必要な物質を調達し、生存し、エネルギーや物質循環をしています。人間的視点から見れば、彼らの作った有機物や鉱物など(硫黄やマンガン、鉄、あるいはメタン、石油?など)も、細菌の活動により、濃縮され蓄積されるようです。
質問:嫌気性細菌にとって酸素は猛毒で生体の全てを酸化し死を誘導すると書いてあったのですが、人間の体内には嫌気性細菌の方が多いと授業で聞きましたが大丈夫なのですか?消化器系の酸素濃度は、呼吸器系と異なり、とても低いです。そのため細菌に限らず消化管内の寄生虫でも嫌気性呼吸をするものが多いです。文献的には、大気の酸素濃度(Po2)は145㎜Hg(21%O2),健康な肺胞の酸素濃度は100~110㎜Hg、健康な大腸の酸素濃度は10㎜Hg(1.4%O2)以下と報告されています。
質問:①深海熱水噴出孔周辺は、生物活動が活発とありましたが水温は何度で、②好熱性を示す細菌は何度まで耐えられるのですか?③熱水に含まれる各種の化学物質にはどのようなものがありますか?テルモプラズマ綱に細胞壁が無く、細胞融合性を持つことの利点は何ですか?①熱水噴出孔はマグマに通じているので高温です。噴出孔から噴出する水温は400℃にも達しますが、熱水噴出孔がある深海の水温は2℃くらいです。 深海の高い水圧によりこの高温でも水は液体のままで沸騰しないようです。②好熱菌は通常、100℃以上でも生育します。古細菌では122℃での生育報告があります。③基本的には、硫黄、鉄、リン、炭酸塩、一酸化炭素、窒素などでしょう。噴出孔の周りに生物が存在すれは有機物ができます。④真核細胞は、原核細胞と異なり細胞壁をもちません。細胞膜が嵌入していろいろな異物を取り込み、時に共生したり、細胞内小器官に変質させたりすることが出来ます。また細胞融合して、細胞が巨大化することは単細胞の活動にとって有利です。原核生物に比べ原生生物のサイズは容積で500~1000倍くらい大きいです。
質問:太古の海は200℃超とあり生命の誕生は熱水噴出孔と書かれていましたが、現在のような水温に下がってからできた熱水噴出孔なのか、太古の海で200℃の中にできた熱水噴出孔なのか、どちらの熱水噴出孔で生命は誕生したのですか?もしくは海全体が熱水噴出孔のような状態だったのですか?46億年前に誕生した地球の表面はマグマの海(1,000℃という高温のマグマ・オーシャン)に覆われていました。その約1億年後、地球が冷え始めたので、空の水蒸気が雨となって激しく降り始め、豪雨の後に地球に海が生まれました。原始海洋は200℃以下となり、温度は150~200℃でした。しかし、40億年前の海でも、大気中の炭酸ガスの温室効果?で、温度は100℃を超えていたと考えられます。しかし、月は現在の1/10くらいの近距離にいたので、10mを超える海水の満ち引きがあり、海水はかき混ぜられ、揺れ動いていたと思われます。このエネルギーと海にある化学物資の反応で生命が誕生したという考えがあります。隕石から来たという、生命の起源に関する別の考えもありますが・・・・
理解できなかったこと:①クロマトフォアと②クロロゾームに関して質問です。クロマトフォアの由来が細胞膜であるのに対して、クロロゾームは由来が細胞膜でないならいったい何からどのようにできたのでしょうか?その材料か構造に違いはあるのでしょうか?
①紅色細菌のクロマトフォアは光合成に必要な成分を多くもった細胞膜小胞で脂質二重膜です。多くの紅色細菌は嫌気状態で,細胞膜のくびれ込みに由来する細胞内膜系(intra-cytoplasmic membranes)を形成します。この膜小胞には光合成反応中心,アンテナ色素タンパク質複合体,電子伝達系成分、ATP合成酵素などの光合成初期反応に必要な成分が含まれています。他方、②緑色光合成細菌のクロロゾームは主に糖脂質(リン脂質)よりなる一層の膜で囲まれた米粒の形をしているアンテナ器官です。バクテリオクロロフィル c 等が会合して棒状になったものを多数内包する脂質一重膜の組織です。包膜である脂質一重膜にはいくつかのタンパク質が存在しクロロソームの形状維持等に関与しています。一重膜ですが両方にグリセロール+リン酸が付くので、外面も内面も親水性になるようです(古細菌の細胞膜型あるいは図のように内面は疎水性のまま?)。真核生物の小胞体、リソゾーム、ゴルジ体などのミクロソーム系はいずれも一重膜?膜構造はもう1度調べる必要があります。

①シアノバクテリアは緑色硫黄細菌と紅色細菌の両方の回路を持っていること、②紅色細菌の一部が、光合成暗反応(カルビン・ベンソン回路)を逆転させクエン酸回路をつくり、酸素が発生するようになったとありましたが、逆転させるの意味がよくわからなかったです。しかも、緑色硫黄細菌があったから変化したのか、紅色細菌のみで変化したのか気になりました。 よろしくお願いいたします。①はシアノバクテリア(藍色細菌)が緑色硫黄細菌の光合成系と紅色細菌の光合成系を合体させた光合成系であるということ。②糖新生系(カルビン・ベンソン回路)と解糖系(クエン酸回路)がちようど鏡像のような関係にあるという意味です。以下に説明します。①シアノバクテリアが2つの細菌の光合成系を合体させたものであるという理由。光合成細菌には、①紅色細菌、緑色硫黄細菌のように、光合成電子伝達系が1種類で、水の酸化能(酸素発生能)を持たないグループと、②シアノバクテリアや真核光合成の葉緑体のように酸素発生型光合成の電子伝達系を持つものがあります。酸素発生型の光化学系は,2つの光化学系(PSI、PSII)の電子の流れがシトクロム複合体を通して繋がれています。①PSIは2つの蛋白質より成るヘテロ二量体構造をもち,アンテナ色素系(光を受け電子を発する)と反応中心(電子伝達)が一体になったアンテナ・反応中心複合体です。ヘリオバクテリアや緑色硫黄細菌にみられる鉄硫黄クラスター型光化学系(PSI型光化学系)は類似し,両者は同一祖先型から派生したものと考えられます。②PSIIは2つの蛋白質より成る構造ですが、この構造は,紅色硫黄細菌や緑色糸状細菌にみられるフェオフィチン-キノン型光化学系(PSII型光化学系)に類似し,両者は共通の祖先型から派生したものと考えられます。従って、酸素発生型の光化学系(シアノバクテリア)の基本構造は,光合成細菌にみられる2つの光化学系(PSI:緑色硫黄細菌とPSII:紅色硫黄細菌)を併せもつことにより成り立っています。②光合成系(シアノバクテリア、葉緑体)と解糖系(αプロテオバクテリア、ミトコンドリア)が逆回転であるという説明。①シアノバクテリアではカルビン・ベンソン回路が回転するので、リブロース(C5)2リン酸に二酸化炭素(CO2)が固定(C6)され、リングリセリン酸(2xC3)となりNADPHからプロトンをもらい、グリセロールアルデヒドリン酸となり、糖(グルコース)と酸素と水を産生する回路です(糖新生系、同化)。他方、②解糖系は、グルコース2リン酸(C6)がピルビン酸(2xC3)となり(2ATP)、ミトコンドリアのクエン酸回路に入り(TCA回路:C2→C6→C5→C4→C4+C2=C6、2TPA)、プロトンポンプを介してシトクロムの電子伝達系へ、プロトンチャネルで輸送して酸素を使って34ATPと二酸化炭素と水を作る回路です。カルビン・ベンソン回路が左周りなら、解糖系+ミトコンドリア(異化)は右周りということになります。鏡像関係にあるという意味で回路の回転が反対になっているという程度の意味です。
質問:原核生物が誕生してから真核単細胞生物が出現するまでには約20億年かかっていますが、①その期間の環境は単細胞生物でいたほうが利点のあるような環境だったのでしょうか?②また多細胞生物は細胞群体のようなものから始まったのですか?③高校生物で真核生物は細菌より古細菌のほうに近いと習ったのですが、原核生物と古細菌の分岐のほうが早いのはなぜですか?①単細胞の方が有利というよりも、原核細胞から真核細胞ができるまでに、20億年かかったということでしょう。原核生物(真正細菌、古細菌)が多様化し、従属栄養で高エネルギー生産が可能になる細菌(好気性従属栄養真正細菌)が出現すること、共生の相手方になる古細菌が大型化し、細胞壁を消失させ、細胞膜を使って、異物を貪食できる能力を開発するのに、やはり時間がかかったのではないでしょうか?酸素濃度の上昇など、環境も少しずつ整ってきた可能性もあります。もちろん、20億年前に突然始まったわけではなく、真核生物になるための試みは何度も起こったはずです。しかし、うまくいかなかったケースは時期尚早だったのではないでしょうか?②現存する多細胞生物の起源は群性化する立襟鞭毛虫(単細胞)であることがゲノムから明らかにされています。体細胞核と生殖細胞核を持った繊毛虫由来の多細胞生物は見つかっていません。でも、どこかにいるかもしれませんね。生物はあらゆることを試みたので、大半は絶滅してしまっています。③古細菌は原核生物です。真正細菌を共生し、真核生物のもとになったと推測されています。真核生物は、理論的には真正細菌と真正細菌の共生or 真正細菌と古細菌の共生 or 古細菌と古細菌の共生のどれかで始まったのでしょう(1次共生)。その後、真核生物と真核生物の共生(2次共生)があったようです。今でも高等生物と細菌の共生はあります。アブラムシのブフネラ菌、植物の内生菌など・・・。
疑問:①最初の生命体とはどのようなもので、どのようにして生まれたのか?②隕石によって飛来したとして、それを証明する手立てはあるのか?どのようにその研究を展開していくのか?①地球上で生命が生まれた場合と、②隕石によって運ばれてきた可能性があります。①の場合は、原始地球の状況を再現し、物理・化学的に有機物を合成し、高分子合成から生命体に持っていく研究で古くからおこなわれています。超高圧、高温、放電条件などで化学合成のアンモニア+CO2→尿素+アクロレイン→尿酸+アンモニア-O2→プリン体にリン酸+りボース(フラノース)が出来れば、核酸が合成できる?RNAそのものに酵素活性があれば、重合して遺伝情報となる?しかしリボゾームがないと、RNAの複製が出来ても、蛋白合成ができない。核酸とアミノ酸が結合するtRNAを作り、RNAから蛋白に置き換える?しかし高次構造蛋白からなるリボゾームがやはり必要・・・。②類似の質問がありました。隕石から最初の生物が来てた場合どうやって証明するのですか? ①隕石中に水の痕跡、②有機体の化石、あるいは③有機物(核酸や蛋白や糖、脂質などの痕跡)が存在していれば、生物がいた可能性が高いと思います。
南極で見つかった火星の隕石?「ALH84001」には、生命の痕跡があるということで話題になりました。その時の根拠は、1) ALH84001に含まれる炭酸塩が生命に適した温度で形成されたこと。2) ALH84001から見つかった有機物は炭素13を比較的少量しか含んでいなくて、生化学反応の痕跡であること。3) 磁性粒子(磁鉄鉱結晶)がバクテリア由来であること。4) ALH84001から見つかった奇妙な構造はバクテリアの化石であること。その後、磁鉄鉱結晶は非生物的に作れることが明らかにされ、この根拠はなくなりました。
現在、世界に存在する細菌の数やウイルスの数は比較的正確に数が推測されていますが、どうやって調べているんですか?僕も気になっていました。1998年、William Whitmanとジョージア大学の彼のチームは、①細菌の生存領域として異なる生息域のタイプ(海洋と他の水生環境、土壌、土壌の地下、空気、動物の内部と葉の表面など)を調べて、②その生息域の数を別々に推定しました。さらに、③生息域は必要に応じてより小さなカテゴリーに分類され(森林土壌対非森林土壌のように)その数が推定されました。④しばしば小カテゴリー別に直接、細菌数が算出されました。⑤直接計算が不可能な場合は、公表されている文献に基づいて推定しました。このようにして地球上に存在する細菌数を推定しました。その結果、地球上の細菌総数は5x1030になりました。
高校までの理解では鳥類から人類が進化したものだと思っていたのだが、人類(哺乳類)は爬虫類のある一種から進化したことがわかった。また、高校生物ではあまり習わなかった五界説についてまだ最初の方であるけれど細菌やウイルスの違いなどについて理解することができた。鳥類が爬虫類から分岐したのは、ほ乳類が爬虫類から分岐したよりもかなり遅いと考えられています。始祖鳥は、現生鳥類の直接的な祖先ではありませんが(系統のとぎれた絶滅種)、ジュラ紀(2億年前~1億4600万年前)の後期、約1億5000万年前のものです。哺乳類の起源は古く、既に三畳紀後期、2億2500万年前には、最初の哺乳類といわれるアデロバシレウスが生息していました。そのルーツは、古生代に繁栄した単弓類 (Synapsida) のうち、キノドン類 (Cynodontia) です。単弓類は、爬虫類の双弓類 (Diapsida) とは石炭紀(約3.5億年~2.9億年前)中期に分岐し、独自の進化をしていました。単弓類は、ペルム紀末の大量絶滅(約2億5100万年前)において壊滅的なダメージを受け、キノドン類などごくわずかな系統のみが三畳紀まで生き延びました。

生物の進化について学んでいるが、そもそも生物が進化しない時の条件はハーディ•バインベルグの法則が成り立っている時と高校生物では習った。しかしハーディ•バインベルグの法則の内の4個(5個のうちのどれを棄却する?)を常に満たすのは困難だと思った。しかしそれでも僕らが今生きている時代で僕は進化というのを目の当たりにしたことがない。この4個の条件を満たしていても特別に進化する時などはあるのか?また進化とは僕らの気づかない内に少しずつ少しずつ起こっているものかなと疑問に思った。吉川先生、解答をお願いします。①ここで問題としている時間は40億年です。突然変異の累積は新種を作り出すには十分すぎる時間です。また、生命史を見れば、最低5回の大絶滅期があり、生物の大半が死滅するという、強烈な選択圧をくぐってきています。もともとハーディ・バインベルグの法則が成り立たないことを前提に考えています。②他方、我々が生きている実時間はせいぜい100年以下です。周りの生物で進化を見るのは難しいかもしれませんが、例えばインフルエンザウイルスの遺伝子を見ればわかるように、流行ごとに遺伝子再集合(リアソート)や突然変異により進化?します。しかし、この場合は自然選択と突然変異がありますね。進化と多様性は、基本的にはゲノムの複製時の遺伝子組み換えと読み間違いによる突然変異を基本としているので、常に少しずつ起こっているとも言えますね。
理解できなかったこと:細菌の構造について質問です。鞭毛が生えている根元の器官ですが、図では細胞膜上にあり、細胞膜・細胞壁を貫通しています。今まで習った範囲ですと、この様な器官は見たことがありません。鞭毛の他にも同じような器官が存在するのでしょうか?線毛も類似の構造です。「線毛は機械の中心から伸び、その長さは細胞の大きさに匹敵することも珍しくない。機械は細胞壁を完全に貫通して存在している。細胞膜に埋もれている回転モーターは新たなサブユニットを追加し、襟の部分は伸びる線毛を外膜の外に誘導する。環状のタンパク質は二層の膜間にある空間を満たしているペプチドグリカン層に機械全体を固定する。」と書かれています。ゲノムをやり取りするには、細胞質同士をつなぐ必要があるので、性線毛は中空で、互いの細胞質に直接開口しているはずです。

疑問:隕石が衝突し、それによって多くの生物が滅んだにも関わらず、なぜ多く生き残ったのがげっ歯類であったのか。本来なら体の大きな爬虫類や両生類などが生き残るのではないのか。また、それらの動物はなにを食べて生き残ってきたのか。約6600万年前、直径約10〜15キロメートルの小惑星が地球に衝突し、恐竜がほぼ絶滅しました。衝突により、約10兆トンの二酸化炭素、1000億トンの一酸化炭素、さらに1000億トンのメタンが一気に放出されたといわれています。また、発生した火災と衝突の衝撃で巻き上げられた塵埃が太陽の光を遮り、全地球規模の気温低下を引き起こし、その結果、植物の光合成が劇的に減少しました。低酸素と低温と少ない食料の中では、恒温(内温性)動物で小型の雑食性で夜行性のげっ歯類に近い哺乳類の祖先が生存に有利となり、生き延び、爬虫類がほろんだ空隙(ニッチ)を埋めるように、繁栄・多様化したと考えられています。大量消費の大型の変温(外温性)動物は、こうした環境には適応できませんでした。
質問:地球上の最初の生命体である細菌の多様性は、化学合成独立栄養細菌から、嫌気性従属栄養細菌、嫌気性光合成独立栄養細菌、好気性従属栄養細菌ときて何故、「好気性光合成独立栄養細菌」が誕生することがなかったのか、その理由があるのか、それとも、存在していたのか、が分かりません。最初は確かに、「嫌気性光合成独立栄養細菌」の方が多かったのですが、シアノバクテリアは酸素を産生する独立栄養の光合成菌細菌です。酸素に耐える能力を持っているので一応好気性と考えられます。しかし、シアノバクテリア(葉緑体)とαプロテオ菌(ミトコンドリア)の両機能を同時に備えた真正細菌が存在したかどうか?真面目に考えていませんでした。インターネットに素晴らしい、専門家の回答がありました。事態はかなり複雑のようです。「シアノバクテリアは、酸素発生型光合成(光化学系Iと光化学系IIが機能する光合成)を営む原核生物として定義されます。ただ、生活様式(代謝系)としては、シアノバクテリアは多様な生物群を包含しており、また、その性質も環境条件によって大きく変化することがあります。1)進化の過程でシアノバクテリアがどのようにして出現したかは不明です(狭義の光合成細菌が起源に関係していることは明らかですが)。シアノバクテリアが出現する以前から地上には低濃度ではあるが酸素が存在していて、酸素を消費する代謝系が発達していたようです。酸素を還元して水に変化させるシトクロム酸化酵素(酸素呼吸の鍵酵素)もその一つです。このような時代から機能している酸素除去系がシアノバクテリアにも備わっています。シアノバクテリアが備えている酸素呼吸の能力が、最初から備わっていたものか、その後の進化の過程で獲得されたものかは分かりません。2)現生のシアノバクテリアには酸素呼吸の機能が備わっているようです。ただ、外部から与えるグルコースなどを取り込んで従属栄養的に増殖できるシアノバクテリアは比較的限られているようです。3)酸素発生の機能(光化学系IIの機能)を備えることによってシアノバクテリアの細胞では、非光合成細胞と比較して、酸素濃度が一万倍程度にも高まったと見積もられています。それに伴って酸素に耐える代謝系が整備されて行ったのだと思います。酸素の存在下で生育する生物と言う意味で、シアノバクテリアは好気性生物です。
理解できなかったこと:細菌の性転換の仕組みについては講義資料より理解できたのですが、細菌の性転換は何を感知して行われるのですか?仮に集団内の性の偏りが感知されて性転換が行われるのだとしたら、♀の個体が減少して新たに♀の個体が必要になるというパターンも考えられると思うのですが、そうした場合は講義資料とは逆に♂から♀に性転換することも可能なのですか?とても面白い質問です。①しかし、生物の世界で雌と雄の比率が1:1に近いケースはむしろ少ないでしょう。2倍体で性染色体をもつ高等生物でも魚類、両生類、爬虫類までは性染色体にかかわらず性転換します。雌雄比はあまり関係しません。環境影響の方が大きいのです。単純な生物では、一般に環境が良いと雌になり、環境が悪化すると雄が出現します。もっと単純な生物では、環境に恵まれれば無性生殖になります。しかし、鳥類と哺乳類くらいは性染色体に従って、約1:1の比で出現します。②おそらく、感知するというより、fプラスミド(f因子)を持つ細菌が性線毛を伸ばし、fプラスミドのない細菌にヒットした時、性線毛が繋がるのでしょう。③fプラスミドを持つ細菌(雄)が2分裂するとき、fプラスミドの入らない方の菌は、雌になるでしょう。またHfrの雄菌は受精させてもfプラスドは崩壊し、受容菌は雌のままです。fプラスミドによる性転換は1万回に1回くらいの頻度と言われています。
理解できなかったこと:深海熱水噴出孔や塩田などはどの生物にとってもかなり厳しい環境なのでかなり大きな進化を遂げないといけないと思うのですが、なぜ古細菌はそのような進化をできて魚などの生物はそのような厳しい環境にも適応できるようにならなかったのですか?とても面白いが、難しい質問です。おそらく、古細菌が分岐した時期(38億年前)は、地球の環境がとても厳しい(高温、高圧、高塩?・・・)状況だったと思います。その後、地球が次第に冷え、雨で薄められる、酸素濃度が上がる・・・など、住みやすい環境になり真正細菌が栄える世界になったのでしょう。極限状況に適応した細菌は少なく、ほとんどが古細菌として進化が止まったのでしょう。魚類などは5億年前に分岐した生物種ですから、極限状況では生存できない細胞から体が出来ています。
質問:1946年にLederberg,J.が大腸菌に雌雄の区別があり、有性生殖を行うことを発見してノーベル賞の受賞したとありましたが、①雌雄の区別があって有性生殖を行えるのは大腸菌だけですか?②また、細菌には環状DNAである、プラスミドが存在していますが、なぜ細菌にはプラスミドが存在しているのですか?③その存在する経緯と雌雄の区別以外の役割は何なのでしょうか?①Fプラスミドには、大腸菌の接合とDNA組換えに関与するF因子があります。しかし、大腸菌以外にもFプラスミドはあります。コレラ菌のP因子,緑膿菌のPF因子などです。②プラスミドは細胞内で複製され、娘細胞に分配される染色体以外のDNA分子の総称です。原核生物の細菌や真核生物で真菌類に属する酵母の細胞質内にも存在します。宿主のゲノムDNAとは独立して自律的に複製を行います。③F因子のようにプラスミドとして細胞質内に存在し、遊離の状態と染色体に組込まれた状態の二つの状態をとることのできる因子をエピソーム (episome) と呼びます。エピソームにはF因子のほか、薬剤抵抗性を支配するR因子 (resistant factor) 、コリシン産生因子、病原性プラスミドなどがあります。プラスミドの由来は、ウイルス同様、明確にはなっていません。
第1回で理解できなかったこととしては、「生命の誕生は40億年前?」のスライドにおいて、研究者らはカナダで見つかった化石は非生物学的理論が当てはまらないので地球最古の生命体の痕跡(有機体)と確信しているとありましたが、その理論とは一体どういうものなのでしょうか。どんな結果から得られた確信なのでしょうか。ケベック州北部のハドソン湾で見つかったこの化石は、①年代的(放射性同位元素による分析?)に37億7千万~42億8千万年前のものと判断されたこと。②化石は、酸化鉄の単繊維とチューブからできており、生物の構造に類似していること(海底の熱水噴出孔のまわりにいる今の微生物が作る構造物に極めて類似)、③酸化鉄が原始細菌により生成されたものと推測されること(炭素化合物と酸化鉄からなる微粒子が見つかった)。④構造物のまわりにはリンを含む鉱物もあったこと。リンは生命の基本構成元素の1つで、死骸が腐敗するときに放出される。⑤今回発見された構造物の分布は無秩序でなく規則的だったこと。当然、異論も出ています。
理解できなかったこと:太陽と地球はほぼ約46億年前に誕生したと言われており、45億年前には海ができ、原始生命体は原核生物(細菌)だというとが分かった。また、約38億年前には真正細菌から古細菌が分岐しました。この時代、生命の歴史の40億年の半分は細菌だけの世界であると分かったがどんな世界だったのだろうと思った。細菌だけの世界といっても、原始地球の隕石やガス、マグマ噴火の灰や高圧の大気、超高熱のマグマと冷えだした地殻、放電や雷、暴風の世界から始まり、水蒸気が雨になり、海が出来、地球から欠けた月が近くにあり、強烈な潮の満ち引きによる大波の中で原始生命である化学合成独立栄養嫌気性細菌が出来た?そのあと独立栄養菌を食べる?あるいは彼らの作った有機物を利用する嫌気性の従属栄養細菌(発酵菌)が出て、弱肉強食の世界の戦いを始めたでしょう。そのうち極限世界に生きる古細菌が分かれ、すみかを広げていきます。最も繁栄した嫌気性の従属栄養細菌が化学合成独立栄養細菌を駆逐し、また、独立栄養菌が利用できる無機物が減ってきたとき、太陽エネルギーを利用して光合成(炭酸ガスと水から糖を作る)をする嫌気性光合成独立栄養細菌がでてきました。その中でシアノバクテリアは光合成で糖とともに酸素を産生するようになりました。海水中の酸素、大気中の酸素濃度が上昇すると今度は、強烈な好気性従属栄養細菌が出現し、主流派となりました。といったストーリーが考えられます。もちろん、いろいろな例外はあります。例えば硝酸菌や硫酸菌のような化学合成好気性菌は、ずいぶん古くからいたようです。

理解したこと: 抗生物質は、人間の手で開発した?ものではなく、細菌の攻撃・防御手段等である事に衝撃を受けました。耐性菌も、もともと持っているDNAが発現をして耐性を持つようになった細菌であるとお聞きして、突然変異では無いんだなと少し安心してしまいました。抗生物質の詳細が、理解出来ました。①突然変異で簡単に耐性遺伝子ができるわけではありません(頻度は少ないのですが、遺伝子の突然変異で耐性が獲得されるケースもあります)が、多くの耐性遺伝子はプラスミドにより運ばれるので、突然耐性菌になる(耐性を獲得する)ことはあります。また②耐性遺伝子には多様性があります(細菌種により点突然変異が起きている)から、問題は複雑です。一度に、数種類の耐性遺伝子を持つプラスミドが伝搬すると、突然多剤耐性菌になります。
理解出来なかったこと:講義の中で錐体細胞は、ヒトでは3種類であり魚類は4種類もつと聞きましたが、進化は必要不必要が大きく関わっているもので夜行性ならば2種類に減るなどは理解できますが、魚類などが4種類以上の錐体細胞を持ち様々な色を認識するのは何故なのでしょうか? 視覚系の進化の中で解説します。魚類でも、ほとんど色覚細胞を持たないものもありますし、昆虫でも同じです。バクテリアロドプシンを基本として、その後、進化の過程で遺伝子が変異、重複を起こし多様化しました。それぞれの系統の生物が環境に合わせて(主に捕食・餌と外敵からの避難のための色の識別に利用)遺伝子を残したり、不活化したりしたのでしょう。基本的には昼行性動物は色覚細胞の種類が多く、夜行性(深海)の動物は色覚細胞の種類が乏しいといえます。集蜜昆虫は花の多様な色を識別するのに必要と思います。魚は熱帯魚のように非常にカラフルな種が多いことや水深による光の透過波長の違いと関係するかもしれません。魚の網膜では、上方(光が多い)を見る部分と下方(光が少ない)を見る部分で波長域の違う錐体細胞が並んでいるという報告もあります。
理解できなかったこと:何のために子孫を残すのか、というのは前々から疑問に思っている事だ。人間が獲得した娯楽的な感情で人生を楽しむのはいいが、他の生物は何をもって一生を良しとするのだろうか。ウイルスもRNA(ゲノム)をわざわざ他の細菌を利用してまで増やしていくが、生き物が増えようとするのは何が目的なのだろう。漠然とした疑問ばかりになってしまいすみません。生き物が生きる目的は、個が一生を生きることとは少し違うかもしれません。般若心経の中に不生不滅、不増不減という節があります。地球が生まれてから現在まで、多くの生物が生まれ滅び、また生まれてきていますが、物質の総量で見れば、ほとんど変わっていないと思います。生き物が増えようとするのは、物質を循環するためであり、物質を循環させるということは、エントロピーを減少させ、永遠の平衡状態(死)に持って行かないためでしょう。地球の鼓動のようなものかもしれません。地球が鼓動をやめるときは、膨張する太陽に飲まれる時か、その前に人間がこの物質の循環を止めてしまう時か?生物の多様性も環境の保全も、エントロピーの減少を保障しようという思いなのではないでしょうか。答えになっていますか?
質問:今回の授業において、最後に出てきた細胞の性転換がよくわかりませんでした。Fプラスミドが性別を決定し、それを細菌同士で与えあうように合成させていくことはわかりますが、もう一つの組み換えの部分はわかりませんでした。①最初の方は、Fプラスミドのコピーが性線毛を通して供与体(雄)から受容体(雌)に移入され、雌から雄になるプロセスです。この場合は、両細菌の遺伝子組み換えは起こりません。性転換といえます。しかし、その後、染色体にFプラスミドが組み込まれることもあるかもしれません。②後の方は、Fプラスミドが細菌の染色体(雄)に組み込まれている場合です。この時は、Fプラスミドとともに供与体(雄)の染色体も一緒に受容体(雌)に移入します。そのため受容体(雌)では、相同遺伝子の組換えが起こります(我々が配偶子を作るときに減数分裂するのと同じです、両親から来た相同染色体間で遺伝子の組み換えが起こります)。この時Fプラスミド部分は分解されてしまうので、受容体細菌(雌)のまま組み換え遺伝子を持つ、全く新しい細菌(雌)になります。
判らなかったこと・疑問に思ったこと:生物の始まりは海中で嫌気性の独立栄養生物からで、次に嫌気性の従属栄養生物が生まれ、嫌気性で独立栄養で光合成をする生物が誕生したということである。ここで疑問が生じた。①最初の嫌気性で独立栄養の生物は海中のどの深さで誕生したのだろうか。仮に海底の熱水噴出孔のような栄養の多い場所で生まれたとすると、深海には光が届かないので、②光合成をする細菌が生まれる頃までに光が届く深さで生きられる能力を獲得したのだろうか。③海中には水圧があるので、深さが浅くなれば深海魚のように膨張して死んでしまうのではないか。④また仮に最初から浅い海の中で誕生したとすると、現に熱水噴出孔の周りなど深海にも細菌が生息していることから、高まる水圧を克服できる能力を獲得し深海まで生息域を拡大したのだろうか。以上、海中で誕生した最初の生物と水圧に関して疑問に思いました。面白い質問です。超高圧と高分子、生物との関係は、わたくしのHPのうち「超高圧と生物」を見てください、いろいろなデータが載っています。④については、好圧細菌はゲノム解析から見ると拡大域を広げていったようです。③ありません。超高圧による細菌破壊力からみて細胞壁の厚いグラム陽性菌は400Mpa(4000気圧、4万メートル)くらいまでは耐えるようですし、芽胞は約1GPaですから水深10万メートルの水圧でも生存します。等浸透圧の多細胞生物とは違います。①②はわかりません。原始地球での最初の生命体は原始海の波打ち際で低分子から高分子化し、RNA合成からはじまったという考えが多いようです。この時期は大気中のガスやマグマの噴火やほこりで太陽光はまだ地球まで十分届かなかったと思われます。
理解できなかったこと:深海熱水噴出孔や塩田などは、どの生物にとってもかなり厳しい環境なのでかなり大きな進化を遂げないといけないと思うのですが、なぜ古細菌はそのような進化をできて魚などの生物はそのような厳しい環境にも適応できるようにならなかったのですか?とても面白いが、難しい質問です。おそらく、古細菌が分岐した時期(38億年前)は、地球の環境がとても厳しい(高温、高圧、高塩?・・・)状況だったと思います。その後、地球が次第に冷え、雨で薄められる、酸素濃度が上がる・・・など、住みやすい環境になり真正細菌が栄える世界になったのでしょう。極限状況に適応した細菌は少なく、ほとんどが古細菌として進化が止まったのでしょう。魚類などは5億年前に分岐した生物種ですから、極限状況では生存できない細胞から体が出来ています。
1つ目は、細菌は、化学合成独立栄養嫌気性→従属栄養嫌気性(発酵)→光合成独立栄養嫌気性→従属栄養好気性と登場しましたが、光合成を行う働きはどこから来たものなのでしょうか。光合成を行うようになったきっかけを教えて下さい。一般には、①原子大気を覆っていたガスや大気中のヘリウムや炭酸ガスが減って、太陽光が豊富に海に届くようになった。②隕石やガスに入っていた化学物質の濃度が化学合成細菌及び嫌気性従属栄養菌の食物連鎖で減少しつつあった。そのため、太陽エネルギーを利用して炭酸ガスと水から糖と酸素をつくる細菌が出現し、繁栄したと考えられています。
2つ目は、吉川先生は、講義の中で人間の体内(消化管内)の細菌は嫌気性が多いと仰いましたが、その理由は好気性になるには酸素が少ないからでしょうか。そうです。それとも、嫌気性に適しているほど硝酸が多いのでしょうか。理由を教えて下さい。消化器系の酸素濃度は、呼吸器系と異なり、低いです。細菌に限らず消化管内の寄生虫でも嫌気性呼吸をするものが多いです。文献的には、大気の酸素濃度(Po2)は145㎜Hg(21%O2), 健康な肺胞の酸素濃度は100~110㎜Hg、健康な大腸の濃度は10㎜Hg以下と報告されています。
ここからは、第2回講義に関する面白かった質問とそれに対する回答集です。色々な質問有難う、勉強になりました。
現代生物進化第2回質問回答まとめ
質問:アメーバの貪食がとてつもない勢いで、白血球などと比べると全く違うという話がありましたが、なぜ人体の防御的機能である白血球系の貪食はアメーバのように激しくないのでしょうか?それくらいの勢いがあった方が風邪やインフルエンザにかかることがなかったのではないのでしょうか?面白い質問です。しかし、多くのアメーバは環境中で単独で生きていくために栄養源として餌を食べます。他方、白血球は血液中の栄養素は細胞膜上のトランスポーター分子などにより取り込むので、栄養源を食べる必要はありません。血液中に侵入した異物(細菌やウイルス)のみを貪食すればいいのです。宿主に寄生し、共生できたアメーバと環境中で独自に生存するアメーバを比較すると、貪食能力や運動能力などが違うかもしれませんね。
質問:古細菌はユーリ、クレン、ナノ、コル・アーケオータの4つに分類されるとありクレン古細菌は好熱菌を中心としたグループで、ユーリ古細菌はメタン菌や高度好塩菌を中心としたグループとありましたが①ナノ古細菌とコル・アーケオータ古細菌はどのような特徴のグループですか?また、ユーリ古細菌は古細菌の8割以上、クレン古細菌は古細菌の2割とありましたが②残りの2つはほとんど存在しないということですか?②ナノ古細菌とコル古細菌は、新しく見つかったグループで、現在、それぞれ数株程度です。純培養は出来ていないようです。①ナノ古細菌(最初の発見は2002年と非常に新しい)は、クレン古細菌に寄生する特別なグループで、細胞の大きさやゲノムサイズが極端に小型化している古細菌です(通常の細菌よりも小さい)。これまで数系統が知られている程度です(好熱菌に入ります)。コル古細菌は1996年にrDNAの解析から、クレン古細菌、ユーリ古細菌と違うグループということになりました。2008年全ゲノム解析が行われました。クレン古細菌に近縁で好熱菌ですが、2018年現在、純培養はできていません。従って、現在わかっている限りでは、ユーリ古細菌とクレン古細菌がほとんどです(合わせて502種、推定950種ですから、同定されている古細菌で400種がユーリ―古細菌、100種がクレン古細菌という割合でしょう。推定種数はその約2倍)。
理解できたこと:二次共生についてです。以前から細胞内共生説は聞いていましたが、単細胞が2回も共生できることは初めて知りました。共生によって器官が増えることは利点だと思いますが、資料のように四重膜までいくと、膜内外の輸送や膜タンパク質の作成が大変だと思いました。膜が通常より多い分、輸送や膜タンパク質が多くなると思うからです。だから動物や植物は二重膜までしかないのでしょうか。ミトコンドリアの外膜は真核細胞の細胞膜、内膜はαプロテオ菌由来と考えていいようです(両方の膜の脂質成分、蛋白とリン脂質の重量比、膜蛋白の種類・機能の違いから)。しかし、植物の葉緑体については外膜も内膜もシアノバクテリア由来という説になりつつあります。3重、4重膜の葉緑体は、細胞内にあっても基本的には食胞内にある細胞外共生(ミドリムシ型)という考え方です。植物の葉緑体は、①真核細胞に取り込まれたのち食胞の膜は消失する。②シアノバクテリアの外膜と内膜が残ったと考えられているようです。僕も初めて知りました。複雑ですね。https://jspp.org/hiroba/q_and_a/detail.html?id=3632
疑問:ミトコンドリアが増殖をする際に内側の膜は自身のDNAから作ればよいと思うのですが、外側の膜の遺伝情報はどうするのでしょうか?宿主の細胞膜のDNAを取り込む仕組みがあるのでしょうか?ミトコンドリアを標識して顕微鏡下で生細胞を観察すると細胞内でミトコンドリアが活発に動き,ミトコンドリア相互の融合と分裂を頻繁に繰り返している様子を観察することができます。細胞内膜の合成は、ほとんど小胞体(ER)で行われ、細胞小器官から出芽した小胞が細胞質を移動し、細胞膜に取り込まれます。このメカニズムで分裂したミトコンドリアを包むのかもしれません。ERでは、細胞質に向いている側の脂質層は常に一定です。膜の合成過程では、小胞体の内腔にて、脂質はまず膜の細胞質側に取り込まれ、フリッパーゼによって一部は細胞外部側へ運ばれます。また、ミトコンドリア膜の融合蛋白(Mitochondria fusion: Mfn)であるMfn2はミトコンドリアのみならず小胞体にも一部局在し,このERのMfn2がミトコンドリアのMfn1またはMfn2と複合体を形成することで,ミトコンドリアとERが結合するとの報告がなされています。

2019年秋①の学生さんの質問から思わぬことがわかりました。
真正細菌と真核生物は細胞膜がエステル型脂質なのに対し、なぜ古細菌だけエーテル型脂質の細胞膜を持っているのでしょうか。エステル型とエーテル型の細胞膜で、なにか違いはあるのですか。大きな違いがあります。①真正細菌と真核生物の細胞膜脂質は、オキソ酸(カルボニル基を持つ脂肪酸)とグリセロール(アルコール)が縮重合(エステル結合R=O-OR)しています。全体は脂肪酸にグリセロール(3位)+リン酸です。反対側の疎水部を中に入れて、脂質2重膜を合成します。②古細菌の細胞膜は、イソブレン(C5)を単位とした炭素鎖からなる脂質とグリセロールが結合したエーテル結合(R-O-R)です。脂質の両側にグリセロール(1位)+リン酸が付くので、両側が親水性になります。③結果として真正細菌と真核細胞の細胞膜は脂質2重膜、古細菌の細胞膜は脂質1重膜です。しかし、例外はあるようです。以前、上に示したように(手書きの図ですが)、細胞膜の脂質2重膜と細胞内オルガネラの1重膜の議論をしました。あの時は、いまいちはっきりしなかったのですが、もう一度、その由来について考えてみます。

理解できなかったこと:また共生の話です。ミトコンドリアの由来のプロテオ細菌が二匹いたとして、その二匹が同時に一つの真核細胞に共生したら、できた細胞の二つのミトコンドリアは別々のDNAを持つのでしょうか。それとも片方しか生き残れないのでしょうか。似た状況は精子と卵子が受精した時、父方のミトコンドリアと母方のミトコンドリアが1つの細胞に入りますが、父方のミトコンドリアはすべて消失してしまい、母方のミトコンドリアしか残りません。また、共生により小器官が増えたら原形質流動はやりにくくなるのではとおもいます。原形質流動は細胞壁があればなおさらです。動物細胞の細胞質ではアクチンケーブルなどの上をダイニンやキネシンなどの移動蛋白にのせて物質輸送が行われるので、ゴルジ体や小胞体のような細胞内膜小器官が増えても問題ないと思います。そうしたらこの生物は体を大きくすれば問題ないと考えます。この繰り返しをすると、いつかは人間のような大きさの単細胞生物ができるのではないでしょうか。なぜ今の地球ではこのような生物が見当たらないのでしょうか。人間サイズのゾウリムシがなぜ存在しないか?もしできたとしても、多細胞動物の餌になってしまったのではないでしょうか?多細胞動物が3胚葉化し、臓器、組織、器官を発達させ、神経系で統合させ弱肉強食を基礎に共進化してきたことを考えると、このゾウリムシは、先カンブリア紀に栄え、カンブリア紀の生物爆発で滅びたエディアカラ生物の巨大生物のようなイメージになるのですが。でも、競争相手のいない極限世界や独立栄養生物しかいない環境では、生きているかもしれませんね。

疑問に思った点:鞭毛虫類のランブル鞭毛虫は、2分裂の無性生殖で増殖し、ミトコンドリアが消失し、代わりにマイトソームを持つようになるとのことですが、マイトソームになった後の無性生殖個体からは一切ミトコンドリアは出てこないのでしょうか。無性生殖は宿主の真核細胞(ジアルジア;ランブル鞭毛虫)のゲノムの複製です。マイトソームはミトコンドリアの遺物で、核に移動したミトコンドリアのゲノムによりコードされています。ミトコンドリアゲノムは、宿主のゲノムに移動してしまったので、再びミトコンドリアとして独立することはありません。また、マイトソームは酸化的リン酸化をしないと調べたら出てきたのですがどのようにエネルギーを得ているのでしょうか。ジアルジアは嫌気性呼吸(酸素を利用しない発酵などの呼吸、ジアルジアは酢酸発酵でATPを作ります)によりエネルギーを得ています。参考資料です。https://www.natureasia.com/ja-jp/natecolevol/interview/contents/4
トリパノソーマの構造のところにて:ヒルは魚類・両生類・爬虫類/吸血性節足動物は爬虫類・鳥類・哺乳類に寄生すると書いてあるが、なぜ爬虫類だけヒルにも吸血性節足動物にも寄生されるのか?教えてください。勘違いです。2つの宿主をもつトリパノソーマは①脊椎動物の血中と②吸血動物の腸管が主な寄生部位です。脊椎動物のうち水生の①魚類・両生類・爬虫類を宿主とするトリパノソーマ種はヒルにより媒介され、陸生脊椎動物の②爬虫類・鳥類・哺乳類を宿主とするトリパノソーマ種は吸血性の節足動物により媒介される。という意味です。おそらく環形動物の水生ヒルに適応した古いトリパノソーマは、水生の魚類、両生類、爬虫類を第2の宿主とした。それに対し、吸血性の節足動物(陸生)に適応した新しいトリパノソーマは、陸生の爬虫類、鳥類、哺乳類を第2の宿主としたのでしょう。ヒルは哺乳動物にも寄生します。ただ哺乳動物にトリパノソーマを媒介するヒルがいないということです。
ゾウリムシはなぜ配偶子も接合子も生じさせないという特殊な分裂をするのですか?
ゾウリムシは無性生殖と有性生殖をおこないます。無性生殖は2分裂によります。体軸方向の前後の部分に分かれるようにして分裂します。有性生殖では、精子や卵子といった配偶子は形成しませんが、生殖核を交換する細胞の接合が行われます。具体的には、①接合に先立ち、大核(体細胞核)が消失します。②残った生殖核である小核が減数分裂を行い、4つの核に分かれます。③このうち3つは消失します。④残った1つがさらに2つに分裂し、⑤このうち1つの核を互いに交換します。その後、⑥それぞれの細胞内の2核が融合することで接合は完了します。しかし、2個体のゾウリムシでは、細胞そのものの融合は行われません。⑦接合後に大核は小核を元に改めて形成されます。
質問:ミトコンドリアや葉緑体が二重膜なのは共生によるものだと高校で習ったのですがもとの細胞をA、共生した細胞を小器官BとするとBの外膜は元々Aの細胞膜ということになり、Bの内膜はB由来ということになるのでしょうか?その通りです。膜はリン脂質と膜蛋白からできていますので、外膜、内膜の特徴は、リン脂質と膜蛋白が真核生物の膜由来か?原核生物由来の膜か?で特徴づけられます。外膜の組成は真核生物の細胞膜と同様に蛋白とリン脂質の重量比がおよそ1:1です。外膜にはポリンという膜蛋白が大量にあり、分子量5000以下の分子が自由に透過できるようなチャネルを形成しており、外膜の進化的起源は真核生物の細胞膜系だと考えられています。他方、内膜はミトコンドリアの機能を担う酸化的リン酸化に関わる呼吸鎖複合体などの酵素群が規則的に配列しています。外膜とは対照的に基本的には不透性で、内外で物質を輸送するためにはそれぞれの物質に対して特異的な輸送体が必要となります。内膜にはミトコンドリアを構成する全蛋白質のおよそ2割が含まれており、蛋白質とリン脂質の重量比は3:1です。内膜の進化的起源は共生細菌の細胞膜を由来としており、内膜に特徴的なリン脂質カルジオリピンの存在がその証拠と考えられています。
質問:トリパノソーマは口になるような器官が図には有りませんが、寄生してどのように宿主から栄養の吸収を行っているのですか?よく気が付きましたね。胞子虫の中でもマラリア原虫、トリパノソーマ、リーシュマニアなどの血中に寄生する原虫は、体表の細胞膜に位置する輸送タンパク質(トランスポーター)を使用して、宿主から大量の栄養素を獲得します。そのため環境中で生きるアメーバや繊毛虫、鞭毛虫のような貪食機能は発達していません。
質問:人間の血液は常に新しく更新されていくのに対して、人間が住血胞子虫に属しているマラリア原虫に感染したとき、マラリア原虫はどうして古い血液と一緒に死ぬことなく、さらにマクロファージなどの免疫系によって排除されることなく、血液中に住み続けることができるのか、が分かりませんでした。①マラリア原虫も古い赤血球と一緒に処理されます。しかし、それ以上に無性生殖します。②原虫もいろいろな手段で生体防御系を潜り抜けます。抗原変異をして免疫系から逃れる。抗原量を増やして免疫系を麻痺させる。シストを作り免疫系に認識させない、免疫系のシグナル経路を阻害する。マクロファージに貪食されてもライソゾームとの融合を阻止する、ファゴライソゾームとなっても分解酵素活性を阻害する・・・などなど複雑です。でも宿主も寄生体に対して遺伝的抵抗性の変異をします。HPの「病原体の科学」第14回に詳しく書いてあります。また、ウイルスの逃げ方は第5回に書いてあります。
質問:藻類で群体を作るイシクラゲやネンジュモなどは数珠繋ぎとなった細胞の中に糸状の藻体があるとの事でしたが、群体となるメリットはあるのでしょうか?あると思います。群体のサイズとエネルギー消費の関係は、正比例ではなく、個体の数が増えれば増えるほど、一個体あたりのエネルギー消費は一定の法則(べき乗則)にそって減っていくようです。群体は単なる個体の集合体ではなく、群体としてエネルギー収支をつけているようです。
分からなかったこと:原生生物でトリパノソーマを出されていましたが①トリパノソーマは具体的にどのような症状を引き起こすんでしょうか?②ハテナについてですが、ハテナは分裂で増殖するんですか?また分裂したときに藻類と原生生物に分かれるとおっしゃつてましたが両方の特徴を持つ(ハテナのまま)増えることはできないんですか?①症状:アフリカトリパノソーマ症は、最初の数か月は無症状。その後、高熱、頭痛、関節痛、肝や脾、リンパ節腫脹。進行すると嗜眠、死亡。アメリカトリパノソーマ症は発熱、眼瞼腫脹、リンパ節炎、その後、肝・脾腫、貧血。死亡の原因のほとんどは心筋炎。②2分裂で増殖します。その時、共生体と宿主の分裂が同調していません。そのため片方の娘細胞(鞭毛虫)には共生体(緑色藻類)が共生します(ハテナのままでいる)。他方の娘細胞は鞭毛虫だけになります。ただ、機会があれば鞭毛虫は緑色藻類を貪食し共生する(ハテナに戻る)ことが出来るようです。
疑問点:今回の授業でアメーバや繊毛虫のような単個細胞は、人間の一つの器官を構成する単個細胞に比べて、その細胞内構造が複雑で、独自に発達した小器官を持っているといいましたが、それは単個細胞として完結して単独で生きる微生物という小さな生物だから、出来ることである。目に見える生物以上のサイズというよりも、多細胞化して、臓器や器官という構造をとると、原生動物の小器官が多細胞の1つの器官に相当するので、個々の細胞だとそのような小器官をそろえることは必要なく、作れないものであり、そのようなことを考えると、原生動物の小器官が多細胞動物の臓器や器官に比べ一概に優秀だといえないのではないですか?このように解釈するなら、それで正しいと思います。またボルボックスのような細胞共生体ではお互いに取り込んでいたエネルギーはどのように共有しているんですか?胚発生の胎児胎盤細胞と内部細胞塊のように、相互にエネルギーや代謝物の需給をしているようになると思います。表面の酸素に触れる部分や栄養素に触れる部分はエネルギー生産が高いでしょう。内部細胞はそのエネルギーや高分子をもらい、違う機能や分化を行っていくのでしょう。
メッセージ:
スライドの中にあった上界、界、門、亜門、網、目これらは分類に使われているものであると思うのですが、なにか基準のようなものがありますか?
①ドメイン(domain)は、界よりも上の、最も高いランクの分類階級です。ゲノムの進化の違い(リボゾーム遺伝子のホモロジーなど)を反映して行われました。その結果、真核生物ドメイン、細菌ドメイン、古細菌ドメインの3つのタクソンが上界にあたります。②真核生物ドメインは、進化の過程で細胞構造(単細胞・多細胞)、栄養の取り方(独立栄養、従属栄養)、動静などに違いが生じた結果4つのグループ(界)に分かれました。原生生物界、植物界、真菌界、動物界。このようなトップダウン方式の基準があります。他方、④ボトムアップ方式があります。動物の種の場合、種を基本単位(交配の可能性、遺伝子の類似性など)として、相互に類縁の深い種どうしを「属」、近縁の属を「科」、近縁の科を「目」、近縁の目を「網」に、近縁の網を「門」に包括されるというように、基本単位のまとまりから順次分類階級を上げていき界につなげる方法があるようです。
疑問点:鞭毛が2本の精子は後方にも泳ぐ事が出来るとありましたが、後方に泳ぐ事が出来るとどんな利点があるのですか?前進のみの車より、前後左右に動ける車の方が便利です。しかし、高等動物を含め多くの雄性配偶子の鞭毛が後方1つ(ユニコンタ)であることを考えると。高等動物の精子は、他の精子より早く卵子にたどり着く競争になったので、直進性の早い方が選ばれたのかもしれません。バイコンタ(前方2鞭毛)のころの雄性配偶子は早さより広い行動範囲を持つことが重要だったかもしれませんね。
メッセージ: この前見ていたドラマでサシガメ由来のシャーガス病についての話がありました。ドラマでは、サシガメの糞を濃縮した液体をあびることでシャーガス病になった、というようなことがありましたが、それだけで病気になるものなのでしょうか。長文失礼しました。私も吸血昆虫という印象だったので糞からの感染?とおもいましたが、調べてみると指摘のようにトリパノソーマが糞中に出るようです。家の土壁、屋根、天井、マットレスなどに生息しているサシガメ類昆虫の糞に含まれるトリパノソーマが粘膜、眼瞼結膜、皮膚刺咬部から体内に侵入することで感染が成立します。就寝中、知らないうちに感染していることがほとんどですので、注意が必要です。また、汚染された飲食物を摂取することによる経口感染、輸血や臓器移植による感染例も報告されています。
高校では一種類だけだと思っていた古細菌がさらに四種類に分けられていること。
この四種類はそれぞれどんな違いがあるのですか?遺伝子の相同性によって分けています。①クレン古細菌は 陸上の温泉などにいる好熱好酸菌で80℃以上の高温を好み海底熱水噴出孔などにいる超好熱菌です。また、②ナノ古細菌は小型でクレン古細菌に寄生しています。③タウム古細菌は中温性の亜硝酸古細菌、④ユーリ古細菌は塩湖や塩田など非常に高い塩濃度を好む高度好塩菌、嫌気環境でメタンを生成するメタン菌、 超好熱菌、好熱好酸菌などを含みます。⑤コル古細菌は、古細菌の中で最も原始的と言われている菌です。最近、古細菌の分類はさらに複雑に分かれています。
質問:ミトコンドリアと葉緑体は独自に増殖するとありましたが、増えすぎては良くないのではないでしょうか?また、どうやってバランスを保って増殖しているのですか?また、ちょうどよい細胞内でのミトコンドリアと葉緑体の個数というものは決まっているのでしょうか?環境によって変化します。ミトコンドリアは細胞のおかれた状況で(①激しい運動や高いエネルギー産生を要求される)数を増やしたり、巨大化します。また、②動物細胞では、低酸素症と無酸素症がミトコンドリアの伸長を引き起こし、③フリーラジカル発生剤は大きなミトコンドリアを誘発します。これは高い酸化ストレスへの適応であることが示唆されました。基本的には大型や多数のミトコンドリアを持つ細胞は酸素消費量が多いと思いますが、低酸素やラジカルストレスへの代償ということもあるようです。センサーは細胞の方にあって、シグナルは細胞からミトコンドリアなどに伝えられると思われます。
理解できなかったことは、原虫を含む原生生物を分類する場合、運動器官によるものが最も一般的なのはなぜなのかということです。運動器官の形態が同じものとして分類したときほかにも共通性は見られるのでしょうか。はいあります。それぞれ系統樹で分岐した時点が異なるために、異なる特性を持っています。例えば、鞭毛虫類は立派な鞭毛で動くだけでなく、2分裂の無性生殖をすること、多くの細胞内小器官をもっています。根足類のアメーバーは高い貪食能を持つこと、体外に排出される時はシストを形成し、シスト内で核分裂を起こし、4核になること、胞子虫類はアピコンプレックスを持つこと、他の原生動物と違い、雌雄の配偶子形成(ガメートゴニー)をすること、繊毛虫類は体細胞核(大核)と生殖細胞核(小核)を持ち、減数分裂し接合すること、消化系、浸透圧調節機構をもつこと等々です。それぞれのグループが独特の共通性を持っています。

核仁と核の違いは何ですか?真核細胞の核膜に包まれた中身を核と言っています。大部分はゲノムDNAで占められています。しかし、この中にRNAを含む領域があり、この部分を核仁(核小体)と言っています。核仁は核の中でも分子密度の高い領域で、リボゾームRNAの転写(DNAからRNAに情報を移し替える)やリボソームの構築が行われる場所です。リボゾームRNAにリボソーム蛋白質が会合して形成されたリボソームは核膜の孔を経て細胞質に運ばれ翻訳(メッセンジャーRNAをたんぱく質に翻訳する)装置として機能します。
質問:藻類は酸素発生をする光合成を行う生物のうち、地上に生息するコケ植物、シダ植物、種子植物を除いたものの総称だと書いてあったのですが、藻類自体は植物ではないのですか?植物の定義が確定しないことが問題です。植物を狭義に共通祖先で地上に生息する①コケ植物、②シダ植物、③種子植物と定義すると、藻類はそれ以外の光合成をする独立栄養生物で酸素を発生するものになります。そうすると藻類は光合成をおこなう酸素発生の独立栄養生物で①~③を除く、生物学的には非常に広い分類になります。真正細菌であるシアノバクテリア(藍藻)から、真核生物で単細胞生物(原生生物)である珪藻、黄緑藻、渦鞭毛藻など、及び多細胞生物の海藻類(紅藻、褐藻、緑藻)など、進化的に全く異なるグループを含んでいます。そうなると「藻類」という呼称は光合成を行うという共通点を持つだけの多様な分類群の総称であり、それ以上の意味を持ちません。しかし、広義に「植物を酸素を発生し、光合成をする独立栄養多細胞真核生物」とすると、今度は、海藻類(紅藻、褐藻、緑藻)などは単純な植物になり、真核細胞である単細胞の原生生物、珪藻、黄緑藻、渦鞭毛藻などが藻類になると思います。
メッセージ: ランブル鞭毛虫はミトコンドリアではなく、代わりに相同と考えられるマイトソームという細胞内小器官が存在し、他には小胞体、ゴルジ体があるがマイトソームは具体的にどんな役割を果たしているのか?同じ鞭毛虫のミトコンドリア相同器官(MRO)であるトリコモナスのヒドロゲノソームでは水素を利用してATPを産生していますが、ジアルジアのマイトソームはATP産生能を失い、代わりに細胞質において解糖系の一部として酢酸からATPを作り出しています。このようにジアルジアのマイトソームは非常に退化しており、残された機能は鉄・硫黄(Fe-S)クラスター合成系だけのようです。鉄・硫黄(Fe-S)クラスターは、電子伝達反応における酸化還元中心として機能するほか, 酵素の触媒中心としての機能, 鉄濃度や酸素濃度などのセンサーとしての役割が知られています。
メッセージ: ガメートゴニーでn+n→2nとなり、スポロゴニー中に減数分裂するとありました。人間は2nの遺伝子で生きていますが、2nではなくnで生きることの利点はなんでしょう?ガメートゴニ―は、無性生殖(クローン増殖、遺伝子組み換えなし)に比べれば、非常に利点は高いですが、2nに比べれば利点はないと思います。Naがガメートゴニ―で雌雄に分かれても、どちらのゲノムもその構造は同じ(Na, Na)です。他の個体の雌雄(Nb,Nb)と接合したとき、はじめてNaNbという2nになります。減数分裂するとNabという新しい個体が出来ます。2nの個体は精子と卵子を作るときに減数分裂するので精子はNab、卵子はNcdになります。受精卵はNabNcdを持つことになります。これが次世代の配偶子を作るときは減数分裂しNabcdとなります。


質問:1胚葉性動物(海綿)はどの部位でもすべての機能を果たすことができるなら、多細胞生物というより単細胞生物が群体をつくったというように捉えることはできませんか?考えてみると、非常にむずかしい指摘です。ボルボックスなどは単細胞の群生?(ボルボックス類も複雑なようです)なのに、海綿は1胚葉の多細胞動物(側生動物)とする理由ですね。定義を見ると、「側生動物(Parazoa)は、生物分類において動物界の一部を構成する亜界(subkingdom)の一つ。「横枝の動物」を意味し、早い段階で動物の多くと進化上分岐したと考えられている。側生動物は肉眼で見える大きさがあり、細胞の分化が見られるという点で、原生動物とは異なる。また、明確な組織や器官を持たない点で、真の多細胞動物(真正後生動物 Eumetazoa)とも異なる。 現存する側生動物は、カイメンなどを含む海綿動物門のみである。」とありますがかなり厳しい説明ですね。ボルボックスでも群生となれば目に見えます。表面側の細胞と内側の細胞は機能が異なり分化して見えますよね。もう少し厳密な差が必要に思います。宿題としましょう。時間があったら調べてみます。

こういう興味ある記述がありました。「われわれは、多細胞生物には多細胞独特の遺伝子があって、そうした遺伝子が多細胞らしさを形作っていると信じてきたが、その考えはどうやら捨てなければならないようだ。非常に多数の遺伝子重複が立襟鞭毛虫(単細胞の原生動物)と(多細胞性)動物の分岐以前に起きていたらしいので、われわれ動物が持つ細胞間情報伝達遺伝子や形態形成遺伝子は単細胞の立襟鞭毛虫にかなり多数存在していると思わねばならない。ひょっとして、立襟鞭毛虫は多細胞生物が単細胞生物に退化したのではないかと思われるほど、遺伝的には多細胞的だ(?)。立襟鞭毛虫ゲノムの全DNA塩基配列の情報は多細胞性の起源と進化について興味深い。立襟鞭毛虫(単細胞の原生動物)と(多細胞性)動物(1胚葉の多細胞性動物:海綿)のゲノム比較から、多細胞化がどのようにして起きたのか、その分子的基礎を解明するための多くの情報が得られるであろう。」現代生物進化の講義で、学生さんとの質疑で、単細胞動物が目玉や腎臓や神経パルスに類似した構造を持つことを理解すると、多細胞化する前に原生動物が多細胞動物の基本遺伝子を作っていたとしてもおかしいことではないと思います。またゲノムサイズだけなら原生動物は細菌レベルのものから肺魚を超すゲノムサイズの持ち主までいるのであるから、遺伝子重複や遺伝子の水平移動は、驚かなくてもいいのかもしれない。ブルーは、原文にはないのですが、吉川が説明をたしました。

ここからは第3回目の講義内容に関する質問です。徐々に質問のレベルが上がってきています。簡単には答えられず、答えを探すためにインターネット使用時間が長くなりつつあります。幸いゴールデンウイークが長期連休だったので、答えが遅れないですみました。
現代生物進化第3回質問回答まとめ
理解できなかった事:動物界の系統区分についてです。高校の生物の授業では、旧口動物の区分の中に扁形動物が所属していたのですが、授業の中で扁形動物は旧口動物に入っていなくて驚きました。三胚葉の動物は旧口動物か、新口動物どちらかに所属するのではないんでしょうか?前口動物(Protostome)は、初期胚に形成された原口がそのまま口となって発生する動物で、原口動物・先口動物・旧口動物ともいい、後口動物に対立します。 広義には扁形動物・輪形動物・腹毛動物・環形動物・軟体動物・節足動物など、多くの動物門が含まれます。刺胞動物などの2胚葉性動物は含まれませんのでその通りです。ただ狭義では、無体腔、偽体腔(扁形動物、線形動物)の動物を除き、真体腔性の動物についてのみ言われることもあります。スライドはこちらの説をとっていたようです。
質問:寄生虫は宿主を介して種の増加・拡大を目的としていることから、潜伏期間は短い方が体外への排出等が早く行われるというメリットがあると考えました。しかしエキノコックスはヒトでの潜伏期間が10年以上と長いという話がありましたが、この潜伏期間の長さによるメリットはあるのでしょうか?ヒトへの潜伏期間の長さはエキノコックスにとってメリットはありません。というのは、ヒトは中間宿主であっていくら原頭節を増やしても、ヒトが終宿主(食肉動物)に食べられない限り意味はありません。しかしエゾヤチネズミのような食物連鎖の間に入る中間宿主では意味があると思います。①中間宿主の中でゆっくり増える(無性生殖する)ことで、1匹のネズミを食べるとキツネは数百匹のエキノコックス幼虫を食べることになる。②中間宿主が長生きをするので、次に生まれた免疫のない子狐が感染ネズミを食べ、処女感染を起こす。③中間宿主(エゾヤチネズミ)が発症する前に生息地域を移動すれば、感染区域を拡張できる。などなど・・・・
質問:①腐生性とはなんですか? ②冬虫夏草は蛾の幼虫に寄生するキノコの一種だと思っていたのですが細かく分類すると違うということでしょうか? ③子嚢菌門の数は他の真菌類に比べてどうして多いのでしょうか?④ヒドラは有性生殖では体の側面に卵巣と精巣を生じ、受精が行われるとありますが、側面にあるのにどの様にして受精することができるのでしょうか?(160文字) ①死んだものを栄養源とする性質です。②冬虫夏草は、昆虫などに寄生するキノコの一種でいいです。正確な生物学的分類では、子嚢菌門、フンタマカビ綱、ボタンタケ目ノムシタケ科ノムシタケ属、シネンシストウチュウカソウとなります。ヒトはサル類から分かれたヒトですが、脊椎動物門、哺乳綱、霊長目、ヒト科、ヒト属、ヒトとなります。③水生から陸生に分岐した時、藻類などと共生し地衣類として栄えたこと、種々の動物に寄生したこと、土壌など環境中に胞子を広げたことが出来たことが大きいと思います。④ヒドラは基本的には、出芽による無性生殖ですが、水温が急激に変化(8℃位)すると異変を感知して雌雄にわかれます。間細胞(幹細胞性)から分化して卵巣と精巣を体表に作り受精卵を雌の体内に残しますが、雌は老化しいずれ死にます。できた受精卵は乾燥に耐えますが孵化する日数は13日~100日とバラツキを示します。これは一度に孵化して悪条件であった場合に絶滅するのを回避するためであるようです。
また。「ヒドラの卵子は卵子限定幹細胞から分化します。精子形成同様に卵子系細胞の増殖で始まり、ネストを形成します。増殖に伴い外胚葉上皮下の数箇所に集合し始めます。ヒドラの卵子形成に特徴的なことは、ひとかたまりの集合細胞のうち、たった一つだけが卵母細胞となり、ほかは全て哺育細胞となることです。卵母細胞ははじめ哺育細胞が放出する細胞質の断片を食作用により取り込み大きくなります。その後、哺育細胞はアポトーシスにより死にますが、卵母細胞は指状の突起を出してこの哺育細胞をも食作用によりとり込みます。とり込む哺育細胞は卵母細胞1個あたり10,000個にもなります。ほとんどの哺育細胞を食べつくすと、卵母細胞は指状の突起を引っ込め、お供え餅のように丸くなります。減数分裂を行い極体を放出します。卵母細胞の真上の外胚葉上皮層に穴があくと、そこから絞り出されるようにして体外に出て、受精可能となります」という記載もありました。

質問:ウェルテルマン肺吸虫の器官では一つの個体に卵巣と精巣の両方があり、回虫のような雌雄異体では雄に精巣、雌に卵巣のように二つの個体に分けられていますが、雌雄異体になったことで多様性や選択圧以外にメリットはありますか?また、ウェルテルマン肺吸虫のほうには雌雄異体のようなメリットはないのですか?雌雄異体のメリットは環境や社会性などの選択圧で選抜された個体が次世代を作るので、より優秀な(適応した)?個体が選ばれることかと思います。デメリットは雌雄が出会わなければ次世代を残せないことです。雌雄同体のメリットは確実に交配できることです。デメリットは自家受粉のように、繰り返すと対立遺伝子がホモザイゴート(近交系マウスのよう)になり、配偶子が減数分裂しても親と子が全く同じ遺伝子構成になってしまう(多様性がなくなる)ことです。
質問1:クマノミなどは群れの中で体の大きい個体が雌雄転換するというお話でしたが、それは彼らが群れ内での自分の体の大きさの順位を視覚的に判断して転換するのですか?それともある一定の大きさになると特別なホルモンなどが出るのでしょうか?視覚的に判断する(喧嘩して勝った方が大きくなる?)ようです。数を理解するのは10匹までで?10匹以下では体の大きさで順位が付き、最も大きい個体が雌になるようです。次は雄。10匹以上を同時に飼育すると見わけがつかず、性分化しないようです(クマノミが理解できる数は10まで)?。質問2:寄生虫は中間宿主と最終宿主をどうやって区別しているのですか?終宿主は寄生すると有性生殖し、受精卵を形成できる種です。中間宿主では子虫のままです。それ以上成長できません。もし最終宿主が別の生物に捕食された場合、その生物に寄生できますか?できません。終宿主は決まっているので、終宿主がほかの生物に食べられると、通常は感染できないで、終宿主とともに消化されてしまうでしょう。卵や原頭節の状態で宿主の胃に入っても死なないのは、特定の生物の消化を防御できる仕組みでもあるのでしょうか?多くはシスト(嚢胞)などに守られて小腸などに来てから脱嚢します。カプセル製剤の薬みたいなものです。
質問:寄生虫にも有性生殖と無性生殖をするものがいますが、講義で①有性生殖のほうが無性生殖より遺伝的多様性が生じるので、種の存続に良いと聞きました。有性生殖を行う際、②雄の個体と雌の個体が必要ですが、ある生物のように③環境の変化合わせて性別が変わる生物もいます。また④雌雄同体の生物もいます。雄と雌が別々の個体として存在する利点はあるのですか?環境に合わせて性別を変化させた方がコストパフォーマンスが良いように感じたのですが、なぜですか?①無性生殖は基本的にクローン増殖なので遺伝子組み換えは起こりません。突然変異以外には遺伝子の多様性が確保できません。②有性生殖でも雌雄同体の受精の場合は、これを繰り返すと対立遺伝子がホモザイゴートになる危険性が高くなります(近交系マウスのようなもの)。③環境に合わせて性転換するのも悪くはありませんが、環境が変化しないと戻し交配となり遺伝子プールが減る危険性があります。④雌雄異体の場合には、効率は良くありませんが、雄間、雌間で環境や社会の選択圧が働き、選択圧に勝つ個体?あるいは選択圧がなければ比較的ランダムに子孫を残すことになるでしょう。というような違いがあります。しかし、魚類・両生類のように一度に多排卵でき受精できる個体群や寄生虫のように中間宿主を持つものは、哺乳類や鳥類(社会性を持ち、世代交代に時間がかかり、少数の子孫しか残せない)とは違う繁殖戦略になっても当然かもしれません。生物の多様性、生態の多様性は、繁殖様式の多様性を生んでいるので、一概にどれが優れているとは言えないかもしれませんね。
質問:エキノコックスはキツネや犬の糞中に虫卵を排出し、その糞が、野菜や草などの食物、水を介して人に経口感染する。人では潜伏期間が10年以上、感染は終生、と講義で教わりましたが、一つの疑問が浮かびました。ブラックジャック本人によるエキノコックス感染です。彼は痛みを感じて、自分で取り除いていましたが、潜伏期間が10年以上あったら、その間、痛みを感じることはないと思われるのですが、どっちが正しいのでしょうか。肝臓自体は痛みを感じることがない臓器ですが、原頭節の無性生殖により病巣が大きくなり肝被膜(肝臓を覆っている被膜)が伸ばされると、内臓痛を感じるようになります。さらに進行すると肝腫大、腹痛、黄疸、肝機能障害などが現れます。また、胆道、脈管などの他臓器に浸潤し、閉塞性黄疸、病巣の中心壊死、病巣感染をきたして重篤となることがあります。末期には腹水や下肢の浮腫が出現します。
海面動物は、組織や器官を持っていないのにどのようにして栄養を摂取しているのですか?カイメンは一般的には体表の穴から水を吸い込み、そこに含まれる微細な有機物微粒子や微生物などを襟細胞などで捕捉し、摂食する。これは海綿の水溝系と呼ばれる濾過摂食のための構造です。しかし、海綿には別種に肉食性海綿があります。肉食性カイメンCarnivorous sponge は、体表の細長い突起で小型の動物を捕まえて食べます。具体的には、体表から細い枝を多数伸ばした形となっており、この部分にある鈎状の骨片により、微少な甲殻類を捕食します。

理解できないこと:幼生と成体で異なる宿主を持つことは生存に不利であり、多くの子孫を残すことを難しくさせるのに、こういう生活環を持っている寄生虫がいるのはなぜですか?①幼生時代に無性生殖できれば、1個の受精卵を取り込んだ中間宿主の中で数百以上の幼生になれる。②中間宿主が飛翔したり、遊泳できれば、自力でなくても中間宿主により、その寄生域を大幅に拡大できる。③より上位の動物に食物連鎖で取り込まれれば、終宿主はより強い動物であるので、安全に安定して受精卵を排出できる。リスクを冒してもメリットも大きいと思います。
わからなかったこと:①なぜ菌類の中でも有性生殖する菌類と無性生殖する菌類がいるのですか?何か違いがあるのですか?②海綿について調べたらの細胞間の結合は緩い と書いてました。波の力で結合が外れてバラバ ラになることはありますか?③線虫類のメスがオスより大きいと書いてまし たがメスがオスより大きい生き物は何か共通点があるのでしょうか?カマキリとか蜘蛛とかアンコウとかがメスの方が大きかったと思うのですが共通点がわかりません。偶然なんでしょうか?人間は大体メスの方が小さいように思うのですがなぜメスの方が大きくないのですか?①無性生殖は基本的にクローン増殖なので、遺伝子組み換えは起きません。そのため多様性は突然変異のみです。有性生殖は減数分裂時に遺伝子組み換えを起こすので容易にゲノムの多様性が起こります。②海綿の種類にもよるようです。強く絞るとばらばらになるものがいます。他方、カイメンは自ら作り出すガラス質の針を、柱と梁のように使って骨格を組み立てていくものもいます。針状構造は骨片です。長さ約150-250 µm,直径4-7µmと非常に微細なガラス質の構造です。側面から見ると、カイメンはこの骨片が柱の様に建ち、テントがポールを支える様に支えます。成長に従いさらに骨片が多数繋げられ、柱と梁構造の骨格が次第に大きくなります。深海性のカイメンの1種、カイロウドウケツの骨格は特に美しいので博物館などで展示されます。③インターネットにそれらしい回答がありました。「寄生虫、昆虫、魚類、両生類などの動物では、産む卵の数が多く、精巣と卵巣の大きさの違いからメスの体が大きいことが多いようです。他方、卵や子供を産む数が少ない場合、オスの体の大きさが大きい場合が多いと思います。哺乳類や鳥類のように卵や子供を生む数が少ない場合、メスは、より生存に有利な子を産めるように、オスを選ぶようになりますし、オスはメスをめぐって競いあうようになります。そうすると、大きいオスのほうが強く、競争に勝ちやすいので、オスが大型化するのだと思います。」なかなか分かりやすい説明と思います。
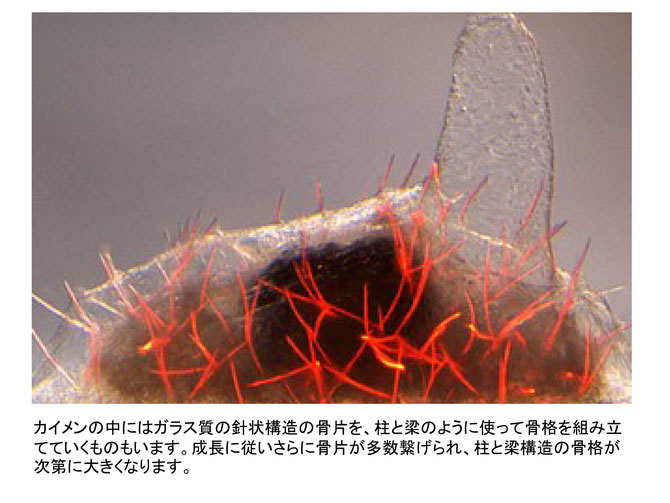
プラナリアは切断・再生後も再生前の記憶があると聞いたことがあるのですが、本当ですか?また、どういう仕組みなのですか?「プラナリアの頭部を切断して、尾部から再生させた個体に、切断前の記憶が残存している可能性」を示唆する実験が、タフツ大学のタル・ショムラット (Tal Shomrat) とマイケル・レヴィン (Michael Levin) によって報告されています。実験の概要は以下のようです。
①数百というプラナリアの環境(温度、時間、水の種類、餌の種類など)を徹底的に均一化し、訓練もすべて自動化しました。光の影響を避けるために、ざらついた表面のあるペトリ皿に移し、ザラツキの上に餌があると学ぶよう、暗闇で10日間の訓練をしました。②プラナリアが光を避ける習性を逆手に取り、青いLEDライトで餌を照明しました。ざらついた表面に餌があると知らない対照群がペトリ皿の縁から動かなかったのに対し、学習した個体は、光を避ける習性があるにもかかわらず、ザラツキを頼りに餌へと到達しました。③記憶が最後の訓練から14日間は持続することを確認しました。その後プラナリアを咽頭前で半分に切断し、後部由来で頭部まで再生した個体(切断から10~14日後)に長期記憶が残されているかどうかを、餌への到達時間を対照群と比べることで判断しました。
④結果は常識的。訓練を施された頭部再生個体と、記憶訓練なしの対照群を比べると、ざらついた表面上にある餌への到達時間はさほど変わりませんでした(ランダムに到達)。⑤しかし、頭部再生個体は、対照群と比べ「再訓練」で学ぶ速度が格段に早く、一度ざらついた表面にある餌を食べさせただけで、頭部再生個体はあたかも訓練を思い出したかのように行動しました。⑥この結果は、プラナリアの「記憶」は条件反射や感作によるものではなく、中枢神経の関与を示唆していると同時に、記憶は脳だけにとどまらないことを示しています。⑦学習により発現した体細胞(体性幹細胞)内の遺伝子(エピゲネシス)が、切断後にエピゲネティックな変化を含んだ中枢神経を再生し、これが「記憶」となる可能性があるかも?https://wired.jp/2013/08/08/memories-of-planaria/
質問:光合成細菌と真菌類を交配して植物と動物のどちらでも繁殖できるみたいなことはできないのですか。真核生物の原生生物同士ですがミドリムシ(ユーグレナ:鞭毛虫と藻類)は、動物と植物のどちらでも生きられます。しかし細菌(原核生物)と真菌類(真核生物)を交配?することは、ゲノムの相同性が低すぎて不可能です。ただし、細菌と真菌の細胞を細胞融合したり、真菌に細菌が共生することはあり得るかもしれません。地衣類は子嚢菌の菌糸に藻類(光合成生物)が共生したものです。菌根菌あるいは内生菌は植物(光合成)に従属栄養の真菌が共生したものです。さらに真菌に寄生する従属栄養植物(もはや光合成できないので植物とは言えない?)もいます。生態系は複雑です。http://bsj.or.jp/jpn/general/bsj-review/BSJ-Review5C.pdf
ツボカビ門のネオカリマスティクス目の真菌は牛や馬にどんな影響を与えるのか?消化を助けています。現在はツボカビ門から独立してネオカリマスティクス門が提唱されています。この真菌類は、絶対嫌気性で主に反芻動物の胃や盲腸などの消化器に存在するルーメン真菌です。反芻動物の胃の中では運動性のある胞子(zoospore)の形で存在し、鞭毛を持っています。ネオカリマスティクスは草食動物の腸内細菌から遺伝子の水平伝播によって獲得したキシラナーゼ(キシランをキシロースに分解する酵素。植物の細胞壁の主要成分の1つであるヘミセルロースを分解することが出来る)やその他のグルカナーゼ(グルコースで構成される多糖であるグルカンを加水分解してグルコオリゴ糖またはグルコースを生成する)といった糖分解酵素を持っています。この酵素を利用して難消化性の繊維が優占する環境下で生活しており、宿主となっている草食動物の消化補助に寄与しています。
疑問点:真菌類は動物寄生種は少なく、大部分が植物寄生種だと習いました。また、動物に寄生する多くの寄生虫も存在するとのことでした。①動物に寄生するタイプと植物に寄生するタイプではどのような違いがあるのでしょう?また、②真菌と寄生虫でも同じような種類の宿主に寄生するタイプでは共通点もあるのでしょうか?①真菌類で動物に寄生するものと植物に寄生するもので違いはあるか?一概には言えません。基本的にはどちらも寄生した宿主から有機物をもらう(生きている場合でも、死体でも)という共通性があります(従属栄養)。しかし内生菌や菌根菌のように真菌のあるものは植物から栄養をもらい、植物に土壌中のミネラル分を与えるものや、植物中に入って生体防御系を刺激するような相互利益の共生系があります。でもキクイムシと真菌や、反芻動物のセルロース分解性真菌のように、動物と相互利益共生する真菌もいます。また、スポロトリクム症のようにバラに寄生する真菌がヒトに直接感染する(植物由来の人植共通感染症)例もあります。植物に寄生する真菌はベジタリアンというべきで、植物性蛋白や脂質をうまく利用するでしょうし、動物に寄生する真菌は肉食で動物性蛋白や脂質を好むかもしれません。②陸生動物に感染する真菌・寄生虫と水生動物に寄生する真菌・寄生虫を考えてみましょう。当たり前で、水生動物に寄生する真菌や外部寄生虫は水生(ミズカビ、ミスダニ)で陸生動物に寄生する真菌や外部寄生虫は陸生(通常の糸状菌、ダニ)になります。陸生の真菌やダニは好気性ですし、ミズカビやミズダニは浸透圧調節に長けていると思います。しかし、生息環境にかかわらすダニ、真菌としての形態や生態には、それぞれ共通性があります。
質問:水中で生活していた植物が陸上にあがり、進化したのにあわせ、陸生となって一部が陸上生物に寄生したという仮説のお話がありましたが、宿主である植物がいきなり陸生になっても、寄生している菌は、そんなにすぐに対応して進化し、寄生し続けることは可能なことなのですか?宿主が植物、寄生しているのが真菌あるいは細菌の場合を考えてみましょう。植物が陸に上がるといっても、おそらく数千万年かけていると思います。多くは、海水から淡水、淡水から岸部、岸部から陸上へと地殻変動による隆起などをきっかけに、徐々に陸生に適応し、そうした植物が繁茂していったと思います。①植物の世代交代が数年単位の時間がかかるのに対し、細菌の世代交代は数時間、真菌であっても条件が良ければ数カ月単位でしょう。世代交代が早いこと、②ゲノムを複製する際に、一定の比率で転写ミスが起こります(1万~10万塩基に1つくらい)。植物の複製酵素や読み取りミスに対する修正システムは細菌や真菌よりも進化しています。その分、細菌や真菌の方が変化しやすい。③植物と共生している細菌や真菌は、植物から糖などの栄養源(有機物)をもらいますが、逆に土壌中のミネラル(窒素、リンなど)を取り込み植物に供給しています。こうした点を考えると植物よりも細菌や真菌の方が、容易に環境に適応する能力(適応したものが選ばれる機会)がありそうに思います。植物や動物が陸上に上がるより、それに共生・寄生している細菌や真菌が陸生に適応する方が容易であると思います。
質問が2つあります。①講義の中で海綿動物は死なないとあったのですが、生殖などは行わないのでしょうか。いくら死なないと言っても、種を繁栄させる為には生殖は必要ですよね? ②プラナリアは、体を切ってもそこからまた体が再生しますが、その仕組みについて、プラナリアの体内がぎっしり詰まっていること(無体腔動物)と関係があるのですか? ①生殖は行います。無性生殖・有性生殖があります。海綿は、1胚葉性の側生動物門に分類されますが4綱25目138科722属8,766種という多様性があります。特徴は、1)組織や器官を欠き,2)細胞は、何にも分化できる分化全能性(totipotency、原始細胞)をもつ数種類の細胞で構成されています。3)襟細胞をもち,水溝系(aquiferossystem)が発達し、4)成体は固着性で懸濁物を食べます、幼生は浮遊性です。5)内外細胞層は基底膜を欠きますが、6)中膠をもちます(アメーバー様細胞が位置する)。7)体に骨片(炭酸カルシウム,珪酸質および海綿質繊維が主成分)を持っています。海綿の生殖は、1)有性生殖の場合、精子は襟細胞からでき、卵は襟細胞か原始細胞(遊走細胞)からできます。体内受精と体外受精があります。2)無性生殖の場合、出芽(budding体外に出る)あるいは芽球(gemmule)が体内で形成されます。②プラナリアが無体腔であることとは直接関係しないと思います。おそらく、多くのプラナリアの体細胞が分化の全能性を持っている(我々の受精卵のように)ことと、細胞同士が極性(前後、左右、上下)を持つことによると思います。プラナリアの再生実験には以下のような報告があります。①頭に切れ込みを入れて3等分にすれば、3つの頭を持つプラナリアに再生する。②メスを使い100を超える断片になるまで切断したが、その全片が再生して100を超えるプラナリアが再生した。トーマス・ハント・モーガンの実験では、遅延や不完全な再生はあったものの、279に分割された断片から再生した。しかし、咽頭および目の前にあたる部分からは、それらが万能細胞(分化全能性)を持たないため再生出来ない(ウィキペディア概要)。
第3回の時に、3胚葉性動物の体腔について無体腔、偽体腔(擬体腔とも書きます)、真体腔の説明をしました。分類上使われているのですが、今後もこの分類が使われるかは意見があるようです。というのは生物分類からすると例外が多いこと、系統進化から見ると退行性に真体腔から偽体腔に変化した動物がいる可能性があること、ゲノムの相同性の分類からは統一性がないことなどから、この分類方法が意味を持つかどうか疑問視されはじめているようです。

わからなかったこと:無体腔動物、真体腔動物は消化管となる内胚葉の周りに筋肉となる中胚葉が覆っているので、食物が消化管に入ってきたとき筋肉を使い食物を消化管の奥に進ませられると思うのですが、偽体腔動物の場合、内胚葉を中胚葉が覆っていないので奥に進ませられなくないですか。どのようにして食物を消化管の奥に進ませているのですか。考えてみると確かに不思議ですね。体壁の筋層が順次収縮して内容物を後ろに送るか(蠕動)?口(咽頭、食道)で飲み込んで順次後ろに送るか?繊毛や絨毛のようなもので後ろに送るか?いろいろあるかもしれません。
以下の記述がありました。①偽体腔動物(例:線形動物の旋毛虫)の筋肉層の内側は体腔であるが、中胚葉性の覆いがないので偽体腔と考えられる。偽体腔の中には体腔液(hemo‐lymph、ヒトのリンパ液や血液のような体液に相当)が充満している。この体腔液によって虫体器官である食道、中腸等の消化管や生殖器官が取り囲まれている。②偽体腔動物のワムシでは取り込まれた食物は、口に続く咽頭にある咀嚼器ですりつぶされ消化管で吸収される。食物は順次口から取り込まれ、咽頭部に送り込まれ咀嚼される(飲み込み型で咽頭部がポンプの役割も果たしていると思われる)。③しかし、以下のような驚くべき記述もあります。「線虫の消化管では筋層、神経叢のいずれも認められない。そこでは,咽頭の自動的収縮が原動力になって食物が移動する受動的な動きがみられるだけである。このためヒト消化管の高次構造や機能は無脊椎動物では未発達で、脊椎動物の出現にともない確立したという認識が生じてもおかしくない。筆者は、この研究をはじめる以前はそのように考えていた。実際は扁形動物無体腔のプラナリアでも筋層、蠕動動運動がみられるし、脊索動物のホヤに筋層、神経叢が存在しないのは咽頭機能の増大によって退化したものと考えられる。従って上記のような認識はあやまりである。本稿に述べる観察結果は、原始的な散在神経系しかもたないと考えられてきたヒドラ(二胚葉性、刺胞動物)に、ヒト消化管の高次構造や機能の原型がすでに存在することを示した。ヒドラに給餌すると餌を次々に捕獲し口に運んで飲み込む。このくり返しで消化管は伸展し満腹状態になる。この過程は嚥下運動に伴う受動的現象だと考えられてきた。他方、少量の給餌を行なうと、食物塊が消化管内を奥にむかって移動する。この移動は、厳密には食物塊の後方の輪走筋が伸展する一方で、前方(口側)の輪走筋が収縮することによって可能になる。哺乳類の食道が行なう運動で、食道反射とよばれる食物塊を咽頭から胃に運ぶ運動と非常によく似ている。この運動は神経系を欠失させた個体では起こらないことから、神経系が関与する能動的現象だと考えられる。」
http://lifesciencedb.jp/dbsearch/Literature/get_pne_cgpdf.php?year=2007&number=5202&file=ilRnpYWpPLUS4gxZ5YPLUSl1IzqQ==
消化管の蠕動運動は、2胚葉のヒドラに原型があるようです。消化系をまとめてみると
①単細胞原生動物の繊毛虫、アメーバなどは食胞で取り込みライソゾームで消化する。
②多細胞で1胚葉の海綿では、鞭毛や繊毛で起こした水流中の微生物をろ過し、
アメーバ様の細胞が取り込んで消化する。
③2胚葉性(無体腔、内外胚葉と間細胞)動物で刺胞動物のヒドラでは口から取り込
み食道の蠕動運動様活動で後方に送る。散在性神経系
④3胚葉性の無体腔動物プラナリアでも同様に蠕動運動と食道の嚥下運動?肛門を持たない(口と肛門が一緒)。籠状神経系
⑤3胚葉性の偽体腔輪形動物のワムシでは、おもに食道の嚥下運動により食物を後方に送る。食道の嚥下機能と咀嚼機能が亢進したかもしれません。口と肛門を持つ。ワムシでは神経系としては咀嚼嚢の背面に脳神経節があり、ここから全身に末梢神経が走る。
⑥3胚葉性の真体腔動物動物(環形動物以上)では、消化管周囲筋層(縦走筋と輪状筋)と神経叢の連動により蠕動運動で後方に送る、となりそうです。

以下に、単細胞真核生物のゾウリムシの食胞、1胚葉性の側生動物である海綿の消化器系。海綿の構造は立襟鞭毛虫の群生に非常によく似ています。鞭毛と襟のろ過部分と食作用部分とアメーバ様細胞からできています。2胚葉性のヒドラでは、細胞分化とその機能は豊富です。内胚葉の消化系細胞の後ろ側の中膠部には筋線維として縦走と輪状筋が走り、神経細胞の支配で消化器系の蠕動運動の原型活動をしているようです。3胚葉性無体腔動物のプラナリアでは、口と肛門が一緒です。餌は口から取り込み、食道から体中に梯子状に分布する消化管に広がり、残渣は食道を介して口から排出されます。蠕動運動と食道の嚥下運動により送り込まれるようです。ほぼ同様に梯子状神経系が分布しています。3胚葉性偽体腔動物である輪形動物のワムシでは餌は口、食道、胃、腸、肛門を通り、泌尿器とともに総排泄口(クロアカ)から排出されます。主に食道の嚥下、咀嚼運動により飲み込まれ、消化管の絨毛により後方に運ばれるようです。

3胚葉真体腔動物の環形動物では、蠕虫(条虫、吸虫、線虫)に比べて消化管の発達は著しく進んでいます。輪状筋、縦走筋が発達し、循環器系や神経系とも密に関連しています。泌尿器系も環境の浸透圧差のためによく発達しています(個々の体節に独立して存在する)。他方、蠕虫で著しく発達している生殖器系はそれほど発達してはいません。寄生虫では、宿主から容易に栄養を吸収出来きるので消化器系が簡略化し、生殖器系が発達する傾向が顕著です。

第4回の講義の主な質問と回答がまとまりましたのでアップします。
今回は海生の節足動物として甲殻類のエビと陸に上がり空を飛ぶ昆虫の代表として社会性を持つミツバチを対象に講義をしました。いろいろ面白い質問がありました。









第5回の講義です。ついに脊椎動物にはいりました。徐々に身近な動物になってきているので、これまでの動物種よりは類推しやすくなっていると思いますが。人間中心の考え方がしばしば裏切られ、生物の多様性が理解できれば幸いです。






理解出来なかった事:生理食塩水は0.8%であると書いてありましたが、0.9%であると仰る先生もいらっしゃり、どちらが本当かわかりませんでした。すみませんでした。生理食塩水の定義は塩化ナトリウムの濃度です。日本薬局方・処方箋医薬品では「塩化ナトリウムを0.9w/v%(厳密には1Lの水に9gのNaClを)含有する食塩水」を「生理食塩液」と定義しています。他方、「人間の体液は、約0.85%の塩分濃度(生理食塩水と同じ?)」に保たれており、これは、ナトリウムとカリウムによって調節されています。細胞外液の塩分濃度が低くなると骨に存在するナトリウムが放出され、逆に、細胞外液の塩分濃度が高くなるとナトリウムは骨に吸収されることで、体液の塩分濃度を調節しています。塩分濃度は、水に溶けている塩の量です。ここで言う「塩分」とは、塩化ナトリウムだけでなく、硫酸マグネシウムや硫酸カルシウムそして炭酸水素塩などの塩類を含めて考えます。また、「人間の体液の塩分濃度は0.8%、それと同じか薄い濃度が体に負担をかけず、おいしく食べられます」という記載もあります。混乱しますが、定義通り、1L量の水に重さ9gの塩化ナトリウムのみを含むものが生理食塩水です。
質問:化石種には海に住む両生類も存在したとありましたが、その場合腎機能はどうなっていたのですか。海水に住む生物、淡水に住む生物、淡水域から海水に戻った生物のどれに一番近かったのでしょうか。また、この場合皮膚呼吸に依存していたのでしょうか。
①両生類の体は塩分に対する耐性が低く、現存種では海産の種は確認されていません。(汽水域に生息する種はカニクイガエル)。ただし化石種には海に住むものも存在しました(チュレルペトン)。両生類の呼吸に関しては全種、幼体・成体を問わず皮膚呼吸が発達しています。②チュレルペトンは全長約50㎝で前後肢とも6本の指を持ち、足自体はいくぶんほっそりし、肩帯もより堅固になり、かなり陸上生活に適応できるようになり、鰓呼吸も失いました。しかし筋肉のつき方は現在の水生有尾類に似た、歩行よりも泳ぎに向いているものです。③より古いアカントステガ、イクチオステガがすでに肺と腎臓を使っているので、呼吸は鰓でなく皮膚呼吸と肺と思われます。腎機能は海に戻った硬骨魚類と同じではないかと思われますが、完全な化石が出ていないので不明です。
他の記述:チュレルペトンが空気呼吸をしていたとしても、主に浅い海に住んでいました。化石が発見されたロシアのAndreyevka層は、この時代の最寄りの陸地から少なくとも200 km離れていました。この地層の水生植物の化石では、その生息域の塩分濃度が激しく変動したことを示しており、海水がかなり浅いことを示しています。チュレルペトンでは頸骨と肩甲帯が分離しているので、頭を持ち上げることができました。したがって、浅瀬では頭が左右に動くだけ、他の動物に比べてかなりの利点がありました。チュレルペトンの近縁種から派生した後期の陸棲動物は、陸上でこの頭の屈曲を必要とします。しかし、これはおそらく陸上ではなく、浅い海域でそれができるという利点のために進化したのでしょう。 「脊椎動物の生活」という本の中で、Pough、Janis、Heiserは、次のように述べています。鰓蓋骨の喪失及びその後の頭蓋骨と脊柱との間の特殊な関節の獲得(まだ初期の四足動物には出来ていない)という明確な首の発達は、呼吸のために鼻を水から持ち上げることや獲物に噛みつくことに関連している可能性があります。チュレルペトンの6本の指とつま先は、それらのもとになった鰭よりも強かったので、浅い海や汽水を進むのに有利でしたが、その手足は、陸上での広範な使用には、まだ十分強くはなかったようです。

イクチオステガが一番最初の原始的な両生類で、海から陸上に上がり、進化していったと思っていたのですが、イクチオステガはほぼ肺呼吸に頼っていたと書かれていたのに、進化していった両生類が肺呼吸だけでなく、皮膚呼吸にも大きく依存しているのが何故なのかわかりません。退化しているように感じます。そうではないと思います。皮膚呼吸は、陸生になった両生類が魚の鱗から皮膚構造を作り替えることにより可能になったものです。鰓呼吸+鰾(気嚢の原基)は魚類の呼吸法です。酸素濃度の乏しい海水を多量に鰓でろ過し、鰓と浮袋に空気をためて呼吸しました。皮膚呼吸+肺呼吸(両生類)は酸素濃度の高い空気から最大限に酸素を取り込む方式に変えました。魚の鱗は皮膚呼吸には向きません。イクチオステガは、陸に上がったが、皮膚はまだ粘膜状で多量の水分で覆われていて、空気中の酸素を取り込むのが、比較的難しかったと思われます。爬虫類は胸腔(肋骨と横隔膜)を発達させ、肺機能を飛躍的に増加させました、また皮膚は厚く乾燥に耐えるように変え(鱗状)、皮膚呼吸をあまり使わなくなりました。肺機能を浮袋(鰾)由来の気嚢+肺呼吸を利用したのが爬虫類から分かれた鳥類です。酸素貯留による呼吸法としては最も優れているかもしれません。哺乳類はほとんど肺呼吸のみで生きています。瞬間的な酸素消費は別にしても、鳥類(渡り鳥)ほど高い高度で長期間の有酸素呼吸を必要としないからかもしれません。哺乳類の肺を使う呼吸法が最も進化した呼吸法であるかどうかは疑問です。また、サメと名についてるものでも軟骨魚類と硬骨魚類に分かれるのはなぜですか。チョウザメとサメのことでしょうか?チョウザメは硬骨魚類です。サメに似ており、鱗が蝶々のような形なので、チョウザメと呼ばれています。チョウザメには浮袋があり、完全な腎臓があり、鯛やヒラメと同じ鰓を持っています。サメは軟骨魚類で歯以外の骨格は軟骨です。浮袋がなく、浮力は油(肝油)、尿素を蓄積し海水の浸透圧に耐えるので、腎臓が未発達です、また鰓は6対の鰓孔から構成されています。チョウザメは単なる名前の問題で生物分類ではありません。サメもどきくらいでしょう。ただし、チョウザメは古代魚の部類に入る古い魚です。
質問①:ヤツメウナギの抗体を作る際のスプライシングのような過程はどのような酵素、また仕組みによって行われているのでしょうか?転写したRNAを一度何か所か切ってバラバラにし、並べ替えてから翻訳しているのですか?多くの蛋白質はパーツ(ドメイン)で翻訳されてから組み立てられます。いまでも免疫グロブリンのHとL鎖は別の染色体上にコードされています。ヤツメウナギでパーツがバラバラにコードされているなら、特にスプライシングしなくても、読まれた蛋白をつなげればよいと思います。質問②:サケやウナギなどの海水と淡水を行き来する魚はどのようにしてそれぞれの場所で体外の浸透圧との差を調節しているのですか?基本的には、塩類細胞の機能を逆転させているようです。淡水では多くの水を排出し、ミネラルを残す。海水では多くの海水をいれ、過剰のミネラルを排出する。質問③:魚類は体外受精で、卵や精子が体外に出たときそれらは単細胞なので塩類細胞はまだないと思うのですが、どうやって体外との濃度差を調節しているのですか?あまり調整しないようです。浸透圧差により機能停止する前に受精する。受精できなかった配偶子はアポトーシスになるのでしょう。精子は核とミトコンドリアと鞭毛で細胞質はほとんどありません。他方、「精子は鞭毛で、さまざまな環境変化を感知しています。海の魚の多くは体内よりも海水の浸透圧が高いことが、精子の運動開始の合図になります。反対に、淡水魚の場合は体外の方が浸透圧は低いため、精子はそれを感知して運動を始めます。ただし、サケ科の魚類の精子は、浸透圧というよりも、周囲のカリウムイオンの濃度低下をきっかけに動き始めます」。魚類の精子の運動時間は短く、1分以下(サケ、マス)、1分(グッピー)から数分(金魚、タナゴ)ですが、長いものは1時間(カジカ)と多様性があります。従って、長時間の浸透圧調節機構は必要ないようです(ミトコンドリアでのエネルギー消費も非常に大きいので、運動でエネルギーが枯渇するとアポトーシスを起こすのでしょう)。卵についても類似の状況です。魚類が水中に産み出す卵にはその周囲に卵巣腔液(あるいは体腔液)とよばれる体液とほぼ、等張な粘性を帯びた液がまとわりついており,この液が低浸透圧環境での精子の運動を助けたり,卵に誘引したり、卵の受精能の急激な低下を抑えている可能性があります。精子と同様に未受精卵も長時間は外環境に耐えられません。数十分から数時間です。魚類の卵は環境水中に放出されると徐々に受精能を失います。淡水魚の未受精卵が淡水中で受精能を失うおもな原因は,卵膜の卵原形質からの分離と,それにともなう卵門の閉鎖です。淡水魚と海水魚の受精卵では浸透圧調節のため受精膜の仕組みに違いがあるのでしょうか?海生魚類の卵の特徴として,水中で発生する胚を物理的に保護するための硬い卵膜(chorion)を持つ点が挙げられます。魚類の卵膜は蛋白により構成される密な構造を持っています。卵細胞質中には蛋白質,核酸,塩類のほかに,比重の軽い油球(oil droplet)を含み、また多くのビテリン由来の遊離アミノ酸を含み浸透圧差に対応しています。受精卵の胚発生が始まると、イオンポンプをはじめ、種々の浸透圧差への耐性機構が動き出すようです。

質問:両生類が皮膚呼吸にとても頼っているとありました。この時皮膚呼吸について調べると両生類は心臓の構造が発達したために(2心房1心室)酸素と二酸化炭素が同時に入っても可能なために皮膚呼吸出来るとあったのですがよくわからないので皮膚呼吸とはなんですか?皮膚の下に分布する豊富な毛細血管に直接酸素を送り込み、ヘモグロビンに取り込ませる方法です。カエルの場合は、冬眠中かそうでないかで異なり、普通は皮膚呼吸が30~50%程度、冬眠中は皮膚呼吸が70%になるといいます。両生類の中にはプレソドン科ムハイサラマンダー(無肺山椒魚)のように肺を持たない種もいます。
質問:ホックス遺伝子とは一体何でしょうか?昨年の質問にありました。ホックス遺伝子というものがどのようなものかわかりませんでした。13個の配列で何が変わるのですか?ホメオティック遺伝子群 (Homeotic genes、Hox genes)は、脊椎、無脊椎動物に関わらず、多細胞動物の胚発生の初期において発現する遺伝子群で、組織、体形の前後、左右、背腹軸および体節を決定する基本的な遺伝子です。この遺伝子は、胚段階で体節にかかわる構造(たとえば脚、触角、目など)の適切な数量と配置について決定的な役割を果たします。順番が入れ変ったり、突然変異が入ると、頭に足が生えたり、お腹に羽が生えたりします。質問:Hox遺伝子についてです。ウニやナメクジウオよりもマウスの方が高次の生物で複雑な遺伝子を持つと思うのですが、マウスの方がホックス遺伝子が少ないのはなぜですか。少なくありません。説明が不足でした。ホメオボックス遺伝子は、よく似た配列をもつ数個の遺伝子(基本は1~13)がゲノムの中に並び、その働きによって、頭部、胸部、腹部などの異なる構造ができあがります。このように並ぶ複数の遺伝子をホメオボックス遺伝子のクラスター(=集団)と呼びます。1980年代の終わり頃には、ショウジョウバエではクラスターが1つなのに対し、マウスなどの脊椎動物では4つということがわかっていました。脊椎動物の祖先と考えられる頭索動物のナメクジウオのホメオボックス遺伝子は全てがそろっていて、最終的にクラスターは1つということがわかりました。脊椎動物の祖先には、もともと1つのクラスターしかなかったのが、脊椎動物が生まれたのと前後してクラスターは4倍に増え、その結果、余分になった遺伝子が変化して、新しい構造をつくるようになったと考えられます。遺伝子数だけからいえば、ナメクジウオが13個、マウスは39個ありまます。
今回の授業で両生類までの生き物は性染色体が発現しにくく結果的に性転換が容易と教えてもらいましたが、逆を返すとこれらと逆の爬虫類、鳥類、哺乳類といった生物が中生代や新生代で幅広く多様に繁栄できたことと関係がありますか?ゲノムが倍加する特性は、両生類(カエル、サンショウウオ)までです。他方、環境による性転換は爬虫類まで起こります。ゲノムの倍加と性転換が起こらないのは、鳥類と哺乳類ということになります。ゲノムの安定性や性転換の廃止が、鳥類、哺乳類の繁栄と直接関係があるという証拠はありません。しかし、雌雄が1:1で生まれ、交配が比較的制約されない(一夫一婦)方が、理論的には遺伝子の多様性は保証されることになります。哺乳類から赤血球は核がなくなり、そして血小板で止血するようになったといいましたが、なぜ哺乳類だけなのでしょうか?哺乳類で赤血球と血小板に核がなくなったのは事実ですが、赤血球が酸素を運び、血小板(栓球)が止血に働く機能は、他の動物種でも同じです。赤血球が核もリボゾームもなくしたのは、哺乳類の赤血球では酸素の運搬能力を最大限にしようとしたという説が主流です。3回の大絶滅(2億5千万年前、2億年前、6500万年前)を生き延びるには、鳥類のように気嚢もなく、昆虫のように気門もなく、皮膚呼吸もなく、肺しか持たない哺乳類の祖先は、ネズミのように小型で、雑食で、恒温性で、低酸素状態にあっても、どこでも生きられる能力が必要であったことでしょう。酸素運搬能力を最大限にするためにヘモグロビンを入れる袋にしてしまったのではないかと考えられています。胎盤性であることと関係があるのでしょうか?ヒトでも胎生期は有核赤血球ですので、特に胎盤性であることとは関係しないと思います。最後に今回の授業で魚の免疫系の細胞についてトランスポゾンの感染を教えてもらいましたが、これは過程は違えど結果的にはミトコンドリアのような細胞内共生説と同じと考えていいですか?一つの細胞に別の生物の機能が入って、発達する事になる。少し違います。ミトコンドリアの時は、他の生物が丸ごと貪食されて共生したものです。単細胞の真核生物でなければ難しいように思います。類似するケースとしては、宿主に細菌、真菌、原虫などが持続感染している状態の方が近い気がします。尤も共生よりは寄生・感染ですが。トランスポゾンはプラスミドや後で学ぶレトロウイルスやレトロトランスポゾンのような、遺伝子運び屋による遺伝子の導入や遺伝子の転移に関係する出来事です。
講義資料には「ゲノムの重複(倍数体)と遺伝子の重複による冗長性を獲得」とあったのですが、これは例えば「ゲノムが重複するコトで鼻を作るための遺伝子も2個分存在するコトになり、その内の1個が仮に変異などを起こして鼻ではないモノを生む遺伝子になってしまっても、もう1個の遺伝子が正常ならば鼻は作られるので、その生物が死ぬコトはなく、むしろ新しい能力獲得の可能性を得る」という理解で良いのでしょうか。それでいいです。また、この理解でいくと「ゲノムの倍数体化でのゲノム量の過多は進化が袋小路になる危険性」というコトの袋小路の部分が判らなくなってしまいました。やはり、はじめの部分の理解を間違えているのでしょうか。間違えてはいないと思います。しかし限度があるとは思います。①ヒトのゲノムを基本にすると、必要な22000個の遺伝子のコード領域は30億塩基の約2%(6000万塩基)です。ハイギョのゲノムがヒトの30倍だから、900億塩基で、ヒトと類似の蛋白を必要とするとやはり6000万塩基なので、0.06%ということになります。1細胞のゲノムの複製にヒトが50倍の無駄なエネルギーを費やすのに対し、ハイギョは1500倍のエネルギーが必要になります。②ヒトが1組の対立遺伝子を持つのにハイギョは30組の対立遺伝子を持ちます。安全確保にはいいけれども、多くは偽遺伝子となります。それよりはゲノムでなく遺伝子のみを重複する方が、無駄がなく安全確保ができるのでしょう(鳥類や哺乳類はこちらを選んだ)。③脊椎動物の原型であるナメクジウオを1倍体とすると高等動物のゲノムが4倍体ですから、この程度の倍数体が安全性・多様性と複製コストとの釣り合い上適当なのかもしれません。④しかし、今でもハイギョやシーラカンスが生存するのだから、絶滅に近いような突然の環境変化が起こった時には彼らは古細菌のように極限環境に適応できるかもしれませんね。
質問:浸透圧調節の方法がいろいろあることが理解できました。等張の体内から高張の海水に出された卵や精子は、まだ調節にかかわる組織がありませんがどのようにして浸透圧差に適応しているのですか。面白い質問です。類似の質問がありました。質問①:ヤツメウナギの抗体を作る際のスプライシングのような過程はどのような酵素、また仕組みによって行われているのでしょうか?転写したRNAを一度何か所か切ってバラバラにし、並べ替えてから翻訳しているのですか?多くの蛋白質はパーツ(ドメイン)で翻訳されてから組み立てられます。いまでも免疫グロブリンのHとL鎖は別の染色体上にコードされています。ヤツメウナギでパーツがバラバラにコードされているなら、特にスプライシングしなくても、読まれた蛋白をつなげればよいと思います。質問②:サケやウナギなどの海水と淡水を行き来する魚はどのようにしてそれぞれの場所で体外の浸透圧との差を調節しているのですか?基本的には、塩類細胞の機能を逆転させているようです。淡水では多くの水を排出し、ミネラルを残す。海水では多くの海水をいれ、過剰のミネラルを排出する。質問③:魚類は体外受精で、卵や精子が体外に出たときそれらは単細胞なので塩類細胞はまだないと思うのですが、どうやって体外との濃度差を調節しているのですか?あまり調整しないようです。浸透圧差により機能停止する前に受精する。受精できなかった配偶子はアポトーシスになるのでしょう。精子は核とミトコンドリアと鞭毛で細胞質はほとんどありません。他方、「精子は鞭毛で、さまざまな環境変化を感知しています。海の魚の多くは体内よりも海水の浸透圧が高いことが、精子の運動開始の合図になります。反対に、淡水魚の場合は体外の方が浸透圧は低いため、精子はそれを感知して運動を始めます。ただし、サケ科の魚類の精子は、浸透圧というよりも、周囲のカリウムイオンの濃度低下をきっかけに動き始めます」。魚類の精子の運動時間は短く、1分以下(サケ、マス)、1分(グッピー)から数分(金魚、タナゴ)ですが、長いものは1時間(カジカ)と多様性があります。従って、長時間の浸透圧調節機構は必要ないようです(ミトコンドリアでのエネルギー消費も非常に大きいので、運動でエネルギーが枯渇するとアポトーシスを起こすのでしょう)。卵についても類似の状況です。魚類が水中に産み出す卵にはその周囲に卵巣腔液(あるいは体腔液)とよばれる体液とほぼ、等張な粘性を帯びた液がまとわりついており,この液が低浸透圧環境での精子の運動を助けたり,卵に誘引したり、卵の受精能の急激な低下を抑えている可能性があります。精子と同様に未受精卵も長時間は外環境に耐えられません。数十分から数時間です。魚類の卵は環境水中に放出されると徐々に受精能を失います。淡水魚の未受精卵が淡水中で受精能を失うおもな原因は,卵膜の卵原形質からの分離と,それにともなう卵門の閉鎖です。淡水魚と海水魚の受精卵では浸透圧調節のため受精膜の仕組みに違いがあるのでしょうか?海生魚類の卵の特徴として,水中で発生する胚を物理的に保護するための硬い卵膜(chorion)を持つ点が挙げられます。魚類の卵膜は蛋白により構成される密な構造を持っています。卵細胞質中には蛋白質,核酸,塩類のほかに,比重の軽い油球(oil droplet)を含み、また多くのビテリン由来の遊離アミノ酸を含み浸透圧差に対応しています。受精卵の胚発生が始まると、イオンポンプをはじめ、種々の浸透圧差への耐性機構が動き出すようです。
理解出来なかったこと:無顎類は顎が無いけど歯はあると先生はおっしゃっていましたが、顎が無いのに歯を使って餌を食べることが出来る理由は何ですか。また、もし歯を使わないのであれば、吸い込むなどすれば良いような気がしますが、では何の為に歯を持っているのですか。面白い疑問です。円口類(無顎類)の歯は、角質歯(honey teeth)です。脊椎動物の中で最も原始的なヤツメウナギの歯は、オタマジャクシの口唇に見られる歯と同じ角質歯です。角質歯(爪や毛と同じ)は、口腔上皮の角質化によって生じます(通常の歯のようなエナメル質は持ちません)。ヤツメウナギの口の周囲と舌の上にある角質歯は,これで魚に吸着し,体表に孔を開け,血液を吸い栄養を採取します。

脊索動物の咽頭部にある内柱という器官は、どのような働きをしているのですか。内柱は、尾索類・頭索類・円口類の幼生の咽頭腹側正中線上に位置する繊毛と帯状に並んだ分泌線をもつ溝状の構造物で、内胚葉由来の器官です。棘皮動物・半索動物には内柱と相同な器官はみられません。この内柱には大きく二種類の機能があります。①粘液蛋白質を分泌する機能、これは、水中の食物粒を捉える濾過摂食のためのもので、数種類の分泌顆粒を含む特化した細胞群がこれを担っています。もう一つは、②ヨードを集積する機能です。円口類成体および顎口類は内柱と呼ばれる形態学的特徴を持ちませんが、ヨードを集積する甲状腺があり、機能的および発生生物学的観点から内柱の相同器官であると考えられています。実際、円口類の甲状腺は成体への変態過程において幼生の内柱の一部から発生します。このように、内柱は脊索動物の共通祖先が獲得した形質で、脊椎動物への進化の過程で甲状腺へと変化する器官です。

分からなかったことがあります。水中のオタマジャクシや硬骨魚類はアンモニアをそのまま排出しているということですが、たとえ排出時には水に速やかに溶けて拡散されるとしても、アンモニアを代謝せず体にとどめておくのは大丈夫なことなのでしょうか。アンモニアは体に良くないのですが、できても、すぐに鰓から捨てるので大丈夫なようです。以下の記述がありました。「大部分の魚は水生生活と関連して窒素の最終代謝産物をアンモニアの形で排出している。肝臓における蛋白の異化過程や運動時に筋肉で生成されるアンモニアは、直接、血液を介して鯉に運ばれ,外界に排出される。水中で呼吸する魚の血液はCO2分圧が非常に低いことと関係して血液のpHは通常約7.8-8.0と高い。このために血液中のアンモニア(NH3)分圧は外界よりも高くなり,NH3は濃度勾配に従って血液から外界に速やかに拡散する。この際,鯉の最外層と環境との接点のpHはCO2やH+の排出によって血液よりも常に低く保たれており,NH3の拡散がより一層促進される。魚はこの様に水中にいる限りは、全くエネル ギーを使用せずにアンモニアを排出することが可能である。」しかし、硬骨魚類でも、尿素型もいます。多様化していますね。
ここからは、第6回目の講義に関する質疑応答です。
ゲノムの数についての質問です。ゲノムというのは2倍体(2n)生物であればn(半数:染色体の1セット)であり、1倍体(n)の生物(無性生殖する多くの生物)であれば、そのままnになるので、基本的にはゲノムにはサイズ(大きさ=塩基数)の違いはありますが、ゲノム1個、2個という考え方はありません。個体の性格や形が異なるのは遺伝子の数や遺伝情報が違うからであって、ヒトはヒト、ショウジョウバエはショウジョウバエというように同じ種類の生き物だったら全く同じ数のゲノム数を持っているのですか。基本的にはそうです。もし、ゲノムの1個、2個という考え方をとるのであれば、それは倍数体ということになります。2n,3n,4n,8n,16n・・・・ゲノム数の多い少ないはどのような違いがあるのですか。いろいろあります、染色体レベルで起こるとトリソミー(特定の染色体だけが3本になる、ダウン症など)や不妊(XXY, YYX, XXYY・・・)になります。しかし、これはゲノムの数が増えることにはならないかも?ゲノムが2倍など倍数体になると種なしスイカの様に大型になりますが配偶子を作れないといったケースが知られています。しかし、我々自身が祖先のナメクジウオから見れば4倍体なので、全ての高等動物は2nといっているが8nのゲノムになっているという考え方もなりたちますか?我々の精子や卵子は減数分裂し、半数体(n)と考えているが実は4nであるとすると混乱しますね。ハイギョは我々が4倍体とすればおそらく120倍体くらいですが、立派な硬骨魚の肉鰭類の魚ですから、形態的には倍数化しても生殖可能であれば、あまり外形的な変化はないのかもしれません。我々は4倍体で2n,ハイギョは120倍体で2nという考え方になりますか。そんな風に考えてもみませんでした。ありがとう!時間がとれたら考えてみます。

質問:生物の分類の中でほとんどの〇〇類が発生した後に数を増やして、最盛期を迎えてまた次に〇〇類の発生・・・の流れだと聞いたことがあります。しかし、その中で鳥類だけは最盛期を迎えていないとそのときに同時に聞きました。鳥類は実際に最盛期?であったことはないのでしょうか?事実だとした場合なぜなのでしょうか?やや似た質問がありました。中生代に脊椎動物の主流は翼竜と鳥類が占めていたが、中生代終わりには翼竜は絶滅してしまった。とスライドにありましたが、鳥類はどうやって生き延びたんですか? ①翼竜(絶滅)は双弓亜綱ですが翼竜目として独立しています。翼竜と恐竜は三畳紀中期(2.2億年)or それ以前に分岐しました。②始祖鳥(絶滅)は鳥綱の古鳥亜綱に属し、現生鳥類に近縁ですが直接の祖先ではありません(1.4億年前には生存)。③現生鳥類に至る歴史は、以下のように考えられています。メイ・ロン:トロオドンと類縁、中国(ジュラ紀後期、1.6~1.5億年前)、シノヴェナトル:竜盤目で白亜紀前期(1.25億年前)に中国に生息したトロオドン科の獣脚類の属の一つ。ミクロラプトルは竜盤目、獣脚亜目で白亜紀前期(1.4~1億年前)に生存、滑空ないし飛行が可能である特徴を持っています(始祖鳥と同じころに生きていました)。ドロマエオサウルスは爬虫綱、双弓亜綱、竜盤目で、白亜紀後期7650万年前~7480万年前に生存、獣脚類恐竜です。トロオドン(Troodontidae)は、双弓亜綱、竜盤目で、白亜紀後期(約7400万~約6500万年前)北アメリカ大陸に生息し、真鳥類に近縁です。これらの恐竜が原生鳥類と最も近い恐竜の一群であり、羽毛恐竜です。6500万年前の大絶滅を生き延びた現生鳥類の祖先(おそらくトロオドンの末裔が、哺乳類同様、その後の世界で繁栄したのだと思います。従って新生代中期から後、鳥類の最盛期は現在を含んでいると思います。

胎盤には機能は同じなのに大きさや形、構成細胞に著しい違いがあるものが複数種類あることがわかりました。またコウモリが哺乳類であることを初めて知りました。また単細胞生物にも目があり、レンズや網膜、色素膜に対応する細胞内小器官を持った生物がいることに驚きました。本当に驚きますよね!性や鞭毛のような運動器官と色覚・光受容体のロドプシンは細菌の時代から、目や消化器、泌尿器系の原基(細胞内小器官)は単細胞時代から、多細胞になって1胚葉の海綿では立襟鞭毛とアメーバ細胞の消化管、2胚葉の刺胞動物では内、外胚葉に神経細胞、消化管の完成、3胚葉のプラナリアでは再生と記憶能力、甲殻類では循環器系、神経系、消化器系、泌尿器系の確立、魚類で特異免疫の原型、鳥類で完成。哺乳類で胎盤というように積み上げてきたと思います。40億年と5回の大絶滅を乗り越えて、よく繋がり、共存してきたという感じですね。
質問:人は免疫では鳥に、初乳ではウシやウマに、視覚では複眼の虫に、色覚では哺乳類に、 色々な点で他の生物に劣っているので人はかなり下位な存在なのではないでしょうか?人は進化があまりできなかったのでしょうか?いえいえ、7回目にはヒトはなぜヒトなのか?その特性は何かを説明しますよ。
理解できたこと:クラゲには目があるのに脳がないのでは、目に入った刺激はどのように伝わるのか。この問いに関して、クラゲはいわゆる反射で動いていると理解しました。ではその逆ではどうなのでしょうか。プラナリアにはnou-darake遺伝子というものがあり、この遺伝子の発現を抑制することでプラナリアの頭部以外にも脳ができると聞いたことがあります。この脳だらけのプラナリアでは、目に入った刺激はどのように伝わるのでしょうか。脳以外にできた目から刺激は神経系に伝わると思います。ただ本来の脳に比べると貧弱です。入力系はできても出力系に繫がらないと行動学的にはわかりません。ただ、光入力にたいして、局所脳が反応することはイメージングでわかるでしょう。もし出力系に繫がると、非常に混乱した動きになるか?再統合されてうまく機能するか?わかりません(沢山の目を持つ動物も結構いるので)。面白いですね。

理解できなかったこと:生物の進化の過程で、遺伝子の重複や倍化が起きたことを聞きました。それでは、この逆はあるのでしょうか。進化の過程で遺伝子や染色体の一部がなくなることは起こらないのでしょうか。程度の問題ですが、あります。実際、T細胞もB細胞も遺伝子の再編後は、遺伝子(染色体の一部)を切り出して捨ててしまいます。またミトコンドリアのゲノムは、もとのαプロテオ細菌からすれば非常に小さくなっています。ブフネラ菌や環境クラミジアをはじめ寄生体ではゲノムの縮小はよく見られます。
疑問:コウモリが鳥類ではなく哺乳類であることについて哺乳類と鳥類の違いが、生えてる毛が体毛か羽毛かどうかである。この二つの毛はどのような条件下で決まるのですか?由来と構造が違います。魚類の鱗は真皮層にできるハイドロキシアパタイトを含む皮骨(歯に近い)ですが、爬虫類の鱗はケラチンを含む表皮が角化したものです。鳥類の「羽毛」は、爬虫類の鱗に由来し、成分はβケラチンです。羽毛には中央に羽軸が存在し、そこから羽肢が枝分かれして生えています。ほとんどの場合、羽軸は一本です(エミュー等は2本です)。成長しきれば、血液は通いませんが、成長中は、血液が通い、形成されていきます。他方、体毛は、毛根部からほぼ一定の太さで生える「毛」で、表皮に由来する爪や角と同じ由来です。成分のほとんど(約80~90%)はαケラチンです。分岐はしません。生えてしまえば、血液は通いません。それぞれ違いはありますが、爬虫類の鱗と鳥類の羽毛と哺乳類の体毛は比較的近い相同器官といえます。簡単にまとめると、魚の鱗は無顎類と顎口類の中間で、初期に絶滅した板皮類の皮骨に近い?硬骨魚類に鱗として継がれていく(歯に近い構造)が、軟骨魚類では捨てられる。陸に上がった両生類で表皮層が発達し、皮膚となるが、爬虫類は表皮を変形させ鱗を作り、これが鳥類の羽毛、哺乳類の体毛(爪や角に近い)になっていったようです。
質問:生まれたばかりの哺乳類は初乳から、あるいは胎盤経由で抗体を得ているとのことでしたが、①その間の抗体は完全に母親由来のものということになり、母親の一部の抗体生産に何らかの異常があれば子も授乳期間その病気にかかりやすいのでしょうか?そうです。母親が暴露されていない病原体には抗体がないので、その病原体に暴露されれば母親も子供も感染します。尤も新生児よりは母親の方が強い(免疫記憶の交差反応が起こりやすいので)と思います。また、母親が無γグロブリン血症、あるいはある種の抗原に免疫寛容なら、子供には抗体は移行しません、あるいは子供はその抗原には免疫状態になりません。②母親が生産することができる抗体のすべてが初乳に含まれているのでしょうか?その場合子によって免疫にばらつきが出ると思います。それとも初乳に含まれる抗体の種類は動物の種内で一定なのでしょうか?妊娠中に母親が持っている抗体のみが移行します。免疫記憶細胞が移行するわけではないので、母親が経験した抗原すべてに対してではありません。また、移行する抗体の種類(サブクラス)には、動物種によりある程度共通性がありますが、抗原に対する抗体(クローン)は個体(母親)ごとに違います。③授乳期の母親の免疫は抗体を多量生産することで上がっている、もしくは子に抗体を与えることで下がるのですか?母体の血液量と胎児の血液量を考えると、抗体が移行したとしても母体の抗体量にそれほど変化はないでしょう。ただ、抗原に対する新生児の抗体価の方が母親の抗体価よりも高いのが普通です。抗体のような高分子の透過性(能動輸送?)に差があるのかもしれませんが、そのメカニズムはよくわかっていません。面白いですね。
疑問点:ゲノムについてがよくわかりませんでした。ゲノムは「切り離せない生物種情報の基本単位」であり「複雑系で意味を持つ単位」とのことですが、それはどのように解析するのですか?現在のゲノム解析は、生物種の全塩基配列を決定し、相互の種の比較をする方法がとられています。主には系統樹を作成したり、特定の遺伝子がどのように多様化したか、ゲノムの重複が起こったか否か?どの時点で起こったか?などが調べられています。また、ゲノム量の変化は何によってもたらされ、何をもたらすのですか?ゲノム量の増大は冗長性をもたらします。必要最小限の情報発現を求めたもの(ウイルスや寄生体)を除くと、生物は情報発現の冗長性(無駄?)を求めたようです。免疫系や神経系も、新しいものが出来たからといって、古い時代の体制を消さないで積み上げ方式で複雑化しています。プテラノドンは恐竜類ではなかったと思います。すみません、どうしても気になったので…了解しました。定義では、正確には「恐竜」は、系統的に異なる翼竜、魚竜、首長竜などは一切含まない独立した分類群でした。この分類群、すなわち恐竜類はそのもっとも際立った特徴として「直立歩行に適した骨格をもった爬虫類」と呼ぶことができ、ほぼすべて地上棲である、とされているようです。翼竜は4足歩行だったのですね?尤も4足歩行になった恐竜もいるようですが。また、哺乳類でもコウモリのように空を飛んだり、クジラのように海にすむものもあるので、地上棲を定義に入れるのはどうかな?また鳥類は直立歩行に適した骨格を持つ爬虫類の末裔?ですが地上棲ではない?ので恐竜ではない?しかし、下の分類では鳥は恐竜、竜盤類、獣脚類でティラノサウルスと同じグループに入るのですね。生物の分類・定義は難しい。
質問:タコやイカには色覚が無いとスライドにはありますが、タコの場合、周囲の色と同化して擬態しますが、色覚が無いのになぜ周囲の色と同化することが出来るのですか。面白い実験報告がありました。①生存のための一戦略として「背地反応」があります。外の環境に合わせて体色を変化させる現象でカメレオン、タコなどで良く知られています。②これに関与する色素胞への神経、ホルモン支配に関しては多くの研究がありますが、その動物が如何にして外の環境の色を知覚するかに関しては分かっていません。③現在ではタコは行動学、電気生理学、視物質研究から色盲とされています。タコの視神経を切断しても背地反応が観察されるので、眼以外に色覚の仕組みがあると考えられています。④ここに挙げられた研究ではタコの皮膚に眼の視物質と類似の光受容体が存在することを検索しました、その結果1)ノザンブロッティング(RNA断片の検出法)解析により皮膚にロドプシン遺伝子とハイブリダイズ(相補反応)する転写産物を見いだしました。2)ウエスタンブロット(蛋白質の検出法)解析により皮膚にロドプシン抗体と交差反応する蛋白を見いだしました。3)レチナールイメージング法により、タコ皮膚の色素胞にレチナール蛋白の局在を見いだしました。4)タコの皮膚の色素蛋白質はタコロドプシンと似た可逆性の光反応を示しました。⑤これらの結果から、タコの皮膚が吸収極大の異なる3つのタイプに分けられることが分かりました。⑥3タイプの皮膚の細胞の、可溶化液の光照射による吸収スペクトルの変化を測定したところ、タコの眼の視物質と同様に青色光の照射により440nmの吸収が減少し、それに橙色光を照射すると470nmの吸収が回復する変化を得ました。この色素を単離するためタコの皮膚をコール酸ナトリウムで可溶化し、ヒドロキシアパタイトにより分離し、可逆的に光反応する分画を得ました。タコの色覚獲得の分子機構を解明するきっかけになることが期待されています。こうしてみると、タコは目だけでなく、皮膚に光受容体をもち、外界の色を識別し、反応している可能性がありますね。

感想:視覚系の進化によるところでどうしてほかの生物のように人間は複眼を持ったりしなかったのかや色覚数が多くないのかと素朴に疑問に思ってしまいました。人間が優れている部分は脳の発達くらいなのかと思ってしまいました。個々の特性から言えばそうかもしれません。運動能力や腕力では肉食獣に、自力飛翔はコウモリや鳥類のようにいきません、目も鼻も大したことはありません。クジラやイルカのように泳げません・・・、やはり脳でしょう。
質問:コウモリにおいて先に飛行能力を得たとあるがそしてそのあとエコロケーションの能力を得たがコウモリはそれ以前はエコロケーション以外に自分の周囲の状況を知る方法はなかったのかについて気になります。夜行性なので目が良いとは思いますが、ほかの夜行性の動物のように目が発達したりほかの部位が発達したりしなかったなのですか?オオコウモリは夜行性(昼行性もいる)ですが、目が発達し鼻もよく発達しています。立体視も可能です。ルーセットオオコウモリはエコロケーションと目と鼻を使います。小型コウモリはエコロケーションを得意としていますが、目が見えないわけではないし、耳が聞こえないわけではありません。また、チスイコウモリやウオクイコウモリ、果食コウモリのように、どの種もエコロケーションで蚊などを食べるわけではありません。しかし、翼手目(小型コウモリ)は比較的早くにエコロケーション能力を発達させたようです。夜に飛翔する夜行性昆虫などを食べる能力は、無敵だったのでしょう。
授業に関係なくて申し訳ないですが、私は人に質問するような疑問がなかなか見つけられません。春の〇〇先生の授業でも毎回苦労していました。そこで質問なのですが、目の付け所が良くなる方法などありませんか。簡単です。①自分の頭で理解しようとして理解できないこと。②枝葉末節でなく本質的なこと、本質的なことは実はよくわかっていません。解けている問題は、どちらかといえば枝葉末節な問題です。今日くれた質問の中に面白いのがありました。イントロンがゲノムの25%をも占めるというのは、何か大きな意味があるのでしょうか。イントロンはスプライシングにより省かれるので、あまり意味のないようなものだと感じていました。しかし、長年の進化の結果、このようになったのは、何かしらの理由があるのではないでしょうか。そうですね、蛋白合成の結果から見れば、基本的にエキソン以外は情報として意味を持たないのでそういうことになります。しかし、最近イントロンの役割についても見直されているようです。「イントロンは一見無駄に見えるが、①選択的スプライシングや、②エキソン・シャッフリングを可能にし、また、③mRNAを核から運び出す過程や、④、蛋白への翻訳効率などに関わっていることがわかってきた。」と書かれています。またイントロンの切り出し方にも原核生物と真核生物では異なるようでいろいろあるようです。①「時にスプライシングを行う部位・組み合わせが変化し、複数種の成熟mRNAが生成することがある。これを選択的スプライシングと呼び、ひとつの遺伝子から多数の生成物が生じてくることになる。選択的スプライシングによってスプライスバリアント(splice variant)と呼ばれる変異蛋白質が生成されるとあります。」抗体のクラススイッチではIgMからIgGに変わるときは、同じクローンで選択的スプライシング(μを飛ばしγ)になりますね。②進化論の話になりますが、「遺伝子を構成するエキソン間の順列組合せが変わる過程をエキソン・シャッフリングといいます。100〜200アミノ酸残基からなる典型的な蛋白質機能ドメインの場合,1つのエキソンのコードする20〜50アミノ酸残基程度のペプチドモジュールが複数集まって機能ドメインが形成されている。ゲノムにエキソン-イントロン構造を有する真核生物の進化において,ゲノム上にあるさまざまなペプチドモジュール単位をコードするエキソンの順列組合せがエキソン・シャッフリングによって変わることで新しい機能をもったタンパク質の遺伝子が効率的に生まれてきたと考えられている。」.確かにこうした例は生体防御系の分子には沢山あると思います。イントロンが関係するのですかね。イントロンごとトランスポゾンでシャッフルされるような気がしますが。③「mRNAとU snRNAの相違点の一つとして,イントロン配列の有無があげられる。mRNAは,イントロン配列を含んだRNA(mRNA前駆体)として転写され、核内でスプライシングされる。一方,高等真核細胞のU snRNAにはイントロン配列がない。イントロン配列の有無が,mRNAとU snRNAとを識別する特徴である可能性がある。検証するため、U snRNAに人為的にイントロン配列を挿入したキメラRNAを作製し、卵母細胞への顕微注入実験を行った。キメラRNAは核内でスプライシングされ核外へ輸送されたが、スプライシングを経たU snRNAにはPHAXは結合せず,替わりにALY/REFが結合していた。このキメラRNAの核外輸送はU snRNAとしてではなく,mRNA型のTAP-p15経路によって行われていた。.以上の結果は,イントロン配列を持つということが,RNAがmRNAとして認識されるための一つの目印であることを示している。」スプライスされる前にイントロンがあることがmRNAの証拠になるのですね。④アンチセンスRNAとは、蛋白質をコードするメッセンジャーRNA(mRNA)と相補的な配列を持つ、ノンコーディングRNA(ncRNA)の一種です。アンチセンスRNAの機能として、mRNAと相補的に結合してタンパク質への翻訳を阻害することがよく知られています。しかし、マウスで見つかったSINEUPと呼ばれるアンチセンスRNAは、mRNAから蛋白質への翻訳を促進するという逆の作用を持っていました。SINEUPの構造の特徴は、標的mRNAに対する相補的な配列に加え、ゲノムに散在する短い反復配列(SINE因子:Short INterspersed Elements)から転写された配列を持つことです。SINE因子は、機能を持たない「ジャンクDNA」と考えられています。SINEUPは、SINE因子由来のRNAに翻訳促進機能が見つかった初めての例です。その後、アンチセンスRNAによる翻訳促進機構がヒトにも存在する(「FRAM」および「MIRb」と呼ばれる配列)ことが明らかにされました」。なかなか奥の深い質問でした。ありがとう。
Hox遺伝子について、勉強しようと思っていたのですが時間が取れませんでした。夏休みに進化論とHox遺伝子に関する面白い論文を2つ見つけました。せっかくなので和訳してみました。まだわからないところがありますが、全体としてのイメージはつかめました。面白い分野です。次回の授業に要旨を取り込みたいと考えています。
A general scenario of Hoxgene inventory variation among major sarcopterygian lineages
主な肉鰭類におけるHox遺伝子データの多様性に関する包括的シナリオ
Dan Liang, Riga Wu, Jie Geng, Chaolin Wang & Peng Zhang
BMC Evolutionary Biology11, 2011
背景:Hox遺伝子は、後生動物の体型を形作る上で重要な役割を果たすことが知られている。従って、これらの遺伝子の進化のダイナミクスは、進化の多様性のパターンを説明する上で不可欠である。総鰭類と四肢動物の両方を含む現存する肉鰭類の中で、Hox遺伝子とそのクラスターに関する我々の知識は、カエル、鳥、哺乳類などのいくつかのモデル生物に限られており、そこには大きな制限がある。そのため、いくつかの進化的なギャップがまだ存在している。このギャップは、特に、特別な体の形態を持つ群や系統樹の主要な位置を占める群など、Hox遺伝子の変異が肉鰭類の系統に沿ってどのように変化するかについての理解を妨げている。
結果:包括的なPCR調査とゲノムウォーキングによって、6種の肉鰭群すなわち肺魚、アシナシイモリ、サンショウウオ、ヘビ、カメ、ワニのHox遺伝子データを決定した。6種の肉鰭群の代表のそれぞれの可変Hox遺伝子を、人間のHox遺伝子データと比較した。肉鰭群の広範な進化をカバーする代表種の遺伝子をPCR法で調査し、その遺伝子の存在/不在についてさらに検証した。カメ、ワニ、鳥、有胎盤哺乳類は同じ39のHox遺伝子を持っている。HoxD12はヘビ、両生類、肺魚には存在しない。HoxB13は、カエルやアシナシイモリで欠失している。総鰭類、両生類および鱗状の爬虫類は、HoxC3を所有している。HoxC1は、アシナシイモリと総鰭類にのみ存在する。現存する総鰭類のシーラカンスと同様に、肺魚にもHoxA14がある。我々のHox遺伝子変異データは、肺魚-四肢動物、カメ類(鳥、ワニ)、カエルーサンショウウオの関係を支持しており、HoxD12の欠失が四肢の減少に直接には関連していないことを示唆している
結論:我々が新たに決定したHoxデータは、肉鰭動物系統に沿ったHox遺伝子の進化ダイナミクスのより完全なシナリオを提供した。 肢のない、ミミズのようなアシナシイモリとヘビは、彼らが由来する体の形態変化が少ない動物と同様のHox遺伝子データを持っていた。これらの体の形態の変化は、Hox遺伝子数の変化ではなく、他の変更による可能性が高いことを示唆している。さらに、本研究の結果は、これらの動物のHoxクラスター全体の将来のシーケンス研究の基礎を提供した。
Hox遺伝子調査の背景
Hox遺伝子は、ほぼすべての後生動物の体軸の形態を制御するホメオボックスを含む転写因子の大きなファミリーである。ほとんどのHox遺伝子は通常2つのエクソン(エクソン1、2)で構成され、保存された180bpのホメオボックスはエクソン2にある。多くの動物種では、Hox遺伝子は最大15の異なる遺伝子メンバーを含むゲノムクラスターに配置されており、重要なことは、それらが「共線的な様式」で表わすことが出来ることである。前部遺伝子は発生の初期に発現し、胚の前方で発現する。後部遺伝子は発生後期に発現し、胚のより遠位の部分に発現する。これらは、前後軸のパターン化に関与する重要な役割のため、Hoxクラスターの変更は、対応する身体領域の変化として現れる可能性がある。従って、クラスターは後生動物の体形プランの多様化を形成する上で、遺伝的革新の源としての役割を果たしている。
Hox遺伝子は、後生動物の形態学的進化の遺伝的基盤を理解する上で特に高い関心がもたれ、頻繁に進化の文脈で調べられる最初の遺伝子群に含まれている。また、Hoxクラスターはさまざまな動物種を特徴付けている。脊索動物の間で、頭索類のナメクジウオは15種のHox遺伝子を持つ単一の完全なHox遺伝子クラスターを持っている。尾索動物のウニでは、単一のクラスターが二次的に破壊され、ゲノム中に分散している。これらの無脊椎動物の脊索動物とは対照的に、原始的な無顎の脊椎動物(円口類:ヤツメウナギとヌタウナギ)は、おそらく無顎類系統の独立したクラスター重複を通じて、3〜7個のHoxクラスターを持っている。有顎の脊椎動物(顎口類)には、脊椎動物といくつかの特定の系統の進化の初期に発生したゲノム複製事象のいくつかのラウンドから生じる複数のクラスターがある。従って、軟骨魚類には3つまたは4つのクラスターがあり、総鰭類と四肢類には4つのクラスターがあり、条鰭類には最大8個のクラスターが、4倍体サケ科の種には〜14個のクラスターがある。脊椎動物のHoxクラスターの変動は、重複の履歴とそれに続く系統固有の遺伝子欠失を反映し、脊椎動物のゲノム進化の研究のモデルとして有効である。
肉鰭類の系統(条鰭類と四肢動物)では、これまでに調査されたすべてのメンバーが4つのクラスター(HoxA、HoxB、HoxC、HoxD)を持っているが、Hox遺伝子のメンバーの数はグループにより異なっている。哺乳類は39のHox遺伝子を持っている。鳥類については、ニワトリのHoxクラスターのゲノム分析に関する最近の報告により、鳥類は哺乳類に対して同じHox遺伝子データを持つ可能性があると推定されるが、ニワトリのゲノムが不完全であるため、2つの遺伝子(HoxC4とHoxC5)がまだ見つかっていない。カエル(Xenopus tropicalis)とトカゲ(Anolis carolinensis)のインシリコ調査では、両方の種でのHoxC3(哺乳類で失われた)の保持が明らかになった。しかし、無尾両生類では、2つの遺伝子(HoxB13とHoxD12)が欠失したと考えられる。最近の関連した研究では、カエルでのHoxB13の欠失はアシナシイモリでも発生し、カエルでのHoxD12の欠失はサンショウウオ、アシナシイモリやヘビでも発生することがさらに報告されている。ごく最近では、インドネシアのシーラカンス(Latimeria menadoensis)の完全なHoxクラスター(初期分岐の肉鰭類)の配列が決定された。四肢動物系統と比較して、シーラカンスはHoxD13を欠く合計42のHox遺伝子を保有している。そこでは、哺乳類で二次的に失われた4つの遺伝子(HoxC1、HoxC3、HoxB10、およびHoxA14)を保持している。
しかし、いくつかの主要なグループからのデータがまだ欠落しているため、肉鰭類系統に沿ったHox遺伝子およびクラスターに関する我々の知識は、まだ完全ではない。例えば、肺魚がシーラカンスのゲノムと同様、Hox14を保持しHoxD13を欠失している(入手可能なデータはやや不完全ではあるが)かどうかを確認するのは興味深いことである。 一方、「ヘビのような」形態(肢のない状態と細長い体)は、ヘビ、アシナシイモリ、アシナシトカゲなどの両生類や爬虫類のさまざまなグループで繰り返し発生する。「ヘビのような」体の形態は、Hox遺伝子の発現の変化が原因である可能性があることを示唆する報告が発表された。これらの特別なグループ間のHoxクラスターの比較分析は、そのような発現の変化がどのように起こるかについてのさらなるヒントを提供できるはずである。Hoxクラスターのシーケンシングには、これらのグループ(ヘビ、アシナシイモリなど)のHox遺伝子データのフレームワーク調査が必要である。
PCR調査は、さまざまな動物のHox遺伝子の予備的同定にその価値を示している。しかし、Hox遺伝子のPCR調査では、縮重プライマー(アミノ酸配列から 未知の遺伝子をスクリーニングする目的で設計されるプライマー、同じアミノ酸に複数のコドンがあるのでプライマーの種類は多い)の優位なバイアスに遭遇することが多いため、既存の遺伝子の実際の数は過小評価されている。この問題を回避するために、当該Hox遺伝子メンバーを標的とする少なくとも2セットの縮重プライマーを設計した。これにより、増幅が成功する確率が高まった。さらに、可能な場合は、テストされた各グループの広い進化範囲にまたがる少なくとも2つの種を選択した。これらのグループでは、プライマーの好みが異なり、グループのすべての遺伝子が見つかる確率が最大となる。これら2つの戦略を採用することにより、アシナシイモリ、サンショウウオ、ヘビ、カメ、ワニ、および肺魚(シーラカンスを除く総鰭類の外にある現存する唯一の種)のHox遺伝子の包括的なPCR調査を実施した。本研究では、肉鰭類の系統内のHoxクラスター進化のより包括的な理解のために、異なる肉鰭類グループ間のHox遺伝子データ変動の全体像を提示することを目的とした。
結果と考察
調査したそれぞれ6種類の肉鰭類について、いくつかのパラロググループ(PG)を対象としたもの、特定のグループを対象としたもの、および特定の遺伝子メンバーを対象としたものなど、縮重プライマーのさまざまな組み合わせを試し、ホメオボックスをコードする領域またはHox遺伝子のエクソン1を増幅した。最初に「一般的な」縮重プライマーを使用して、次に最初の調査で見つからなかったパラロググループまたは遺伝子メンバー用に設計されたより特異的なプライマーを使用して、数ラウンドのPCRを行った。合計3876のPCRフラグメント(使用するプライマーに応じて80〜165 bpの長さ)をクローニングし、シーケンスした。ライブラリーの構築と6種類の肉鰭類およびその他の関連種のスクリーニング効率に関する詳細情報は、「追加ファイル1:シーケンスされたクローンの統計」にリストとして示した。たまに、ヌクレオチドが1〜3個だけ異なる2つのシーケンスが見つかった。ヌクレオチド変異が同義置換に属し、これらの各配列が複数のクローンに存在する場合、それらは対立遺伝子変異体と見なした。逆に、1つのクローンのみの1つまたは2つのヌクレオチドの非同義置換は、同じフラグメントの他のものではなく、PCRまたはシーケンスエラーと見なし、以降の分析から除外した。
GenBankでの最初のBLAST検索では、調査した動物種に応じて、82.9〜96.5%のシーケンスクローンがHoxフラグメントに属していることが示された。 この結果により、ほとんどの肉鰭類系統にわたって新しく設計されたHox調査プライマーの高い有用性と効率が実証された。蛋白質レベルでの系統解析に基づき、得られたホメオボックス配列の70〜75%を正確なHox遺伝子メンバーに明確に割り当てることができた。Hox2、Hox6-8の蛋白質アラインメントの系統学的シグナルは特に弱かったため、核酸レベルでの系統学的分析を行い、これらのパラロググループのHoxメンバーを区別した。 この戦略を使用して、取得したすべてのHoxフラグメントのオルソロジーを決定することができた。
核酸系統解析に基づく推定Hoxフラグメントの割り当ての信頼性をテストするために、2種の動物(タイワンコブラとバンナアシナシイモリ)を選択した。これらの動物の Hox6-8フラグメントの推定される3 '隣接シーケンスを取得するためにTAIL-PCRベースのゲノムウォーキングを実行した。 新しく得られたシーケンスは、既知の肉鰭類Hox遺伝子と比較され、すべての割り当てが検証された。
異なる動物群のHox遺伝子データ
肺魚
アフリカの肺魚(Protopterus annectens)は、合計42のHox遺伝子を持っていることがわかった。それらはシーラカンスの4つのHoxクラスター遺伝子に対してオーソロガス(相同)である。一般に、アフリカ肺魚のHoxデータはシーラカンス(Latimeria menadoensis)のそれと非常に似ている。たとえば、どちらもHoxC1、HoxC3、HoxB10を所有している。しかし、興味深いことに、アフリカ肺魚にはHoxD12が、シーラカンスにはHoxD13が欠失している。さらに、シーラカンスにはHoxA14遺伝子があり、軟骨魚類にはHoxD14遺伝子がある。これまでに調査されたすべての四肢動物ではHox-14を欠いているので、Hox14メンバーが肺魚類および/または基本的な四肢動物系統(アシナシイモリ)にも存在するのかどうか疑問である。
これを確かめる目的で、Hox14でのみ保存されている84bpフラグメントをアフリカ肺魚とBannaアシナシイモリに対しPCRで調査した。その結果、アフリカ肺魚だけで予想されるサイズのバンドが検出されたことから、Hox14がすべての四肢動物で失われた可能性があることが示唆された。しかし、肺魚のHox14フラグメントはサイズが短いため、PCRではこれ以上の分析はできなかった。従って、次にHox14フラグメントの両端に向かってゲノムウォーキングを行い、肺魚Hox14のエクソン2の完全な配列を得た。そこでヤツメウナギHox14αをアウトグループとして、系統学的分析を行い、肺魚のHox14メンバーがHoxA14として明確に割り当てられた。これまでに調査されたすべての脊椎動物の中で、現存する総鰭類の唯一の2群であるシーラカンスと肺魚だけがHoxA14を持っていた。したがって、HoxA14は、総鰭類のHoxレパートリーの特徴である。
アシナシイモリ
Bannaアシナシイモリ (Ichthyophis bannanicus)では、39のHox遺伝子が検出された。 Hox遺伝子のコアセット(参照として哺乳類を使用)に加えて、Banna アシナシイモリでHoxC1とHoxC3が検出された。アシナシイモリにおけるHoxC1の存在は、分岐したアシナシイモリの代表であるGymnopis multiplicataのHox1ライブラリーをスクリーニングすることによって検証された。 HoxC3はカエル(Xenopus tropicalis)とサンショウウオでも発見されたため、現存する両生類はすべてHoxC3を保持していると思われる。そのため、他のアシナシイモリ種ではHoxC3の調査は行わなかった。さらに、プライマーのさまざまな組み合わせ作成した後、アシナシイモリ3種(I. bannanicus、G. multiplicata、Typhlonectes natans)で分析した結果、HoxD12およびHoxB13は検出されなかった。これは、以前にMannaert et al.によって報告されたアシナシイモリにおけるHoxD12とHoxB13の欠失と一致した。
サンショウウオ
他の動物群に比べサンショウウオのHox遺伝子データの調査はより困難であった。我々は、いくつかの種を試したが、それらのどれにも33を超えるHox遺伝子を検出することが出来なかった。サンショウウオのより統合されたHox遺伝子データを得るために、2種(Batrachuperus tibetanusとPachytriton brevipes)から得られた結果を組み合わせてグループとした。 、チベットサンショウウオ(Batrachuperus tibetanus)のHox遺伝子の最初のPCR調査では、770クローン以上あった。シーケンス後に33のHox遺伝子を特定した。カエルのHox遺伝子と比較して、HoxA1、HoxA3、HoxD3、HoxB6、HoxB7、HoxA10が見つからなかった。不足しているHox遺伝子は別のサンショウウオPachytriton brevipesでさらに調査され、220クローンのシーケンス後に2つのHox遺伝子(HoxA1とHoxA10)が検出された。残りのHox遺伝子の存在を検証するために、それぞれのexon1を検出し、最後にHoxA3、HoxD3、HoxB6、HoxB7の遺伝子が検出された。 Salamandrids およびambystomatids でのHoxB13の存在に関する以前の報告と一致して、より原始的なサンショウウオ(B. tibetanus)でHoxB13を検出できた。以前の調査結果と同様に、調査したすべてのサンショウウオでHoxD12を検出できなかった。サンショウウオは39のHox遺伝子を持ち、そのうちHoxC3を持ち、HoxD12が欠失している。また、他の2つの両生類グループ(アシナシイモリ、カエル)とは異なり、サンショウウオはHoxB13を保有している。
カエル
カエル(Xenopus tropicalis、二倍体のカエル種)のHoxクラスターのゲノム構造は以前に報告されている。HoxB7はシーケンスギャップのために検出されなった。 ただし、HoxB7 mRNAは別のカエル(Xenopus laevis)でクローン化されており、カエルがHoxB7遺伝子を持っていることは明らかである。 総計、カエルには38のHox遺伝子があり、HoxB13とHoxD12は欠失しているが、HoxC3は保持されている。
トカゲとヘビ
Di-Poïet al. は、グリーンアノールトカゲ(Anolis carolinensis)のゲノムデータを分析し、トカゲには40のHox遺伝子があり、さらに哺乳類には存在しないHoxC3遺伝子があることを報告した。ただし、HoxB13とHoxD9は、研究で推定された遺伝子のみであり、いくつかのシーケンスのギャップのために直接検出されなかった。これらのギャップのフランキングシーケンスを再分析し、HoxB13(exon2)およびHoxD9(exon1)にオーソロガスなフラグメントを特定し、トカゲのHox遺伝子データの完全性を改善した。ヘビの場合、中国コブラ(Naja atra)に39のHox遺伝子が検出された。トカゲと同様に、ヘビにもHoxC3遺伝子が存在した。 HoxC3の存在がすべての有鱗動物の特徴であるかどうかを確認するため、他の2種の有鱗動物、ゲコ(Hemidactylus bowringii)とブラインドスキンク(Dibamus bourreti)のHox-3遺伝子をさらに調査した。その結果、これら2種でのHoxC3遺伝子断片が特定された。これらは、全ての有棘動物がHoxC3を所有していることを示唆している。グリーンアノールトカゲでHoxD12が検出されているにもかかわらず、異なるプライマーペアで構築されたHox12ライブラリをスクリーニングしても、中国コブラでHoxD12遺伝子は検出されなかった。 HoxD12の欠失は、最近、コーンスネークで報告されている。中国コブラとコーンスネークの両方ともヘビ類に属しているため、全てのヘビにHoxD12が存在しないかどうかをさらに調査するため、他の2種のヘビでHox12を調査した。ブラインドヘビ(Leptotyphlops blanfordii)とボールパイソン(Python regius)であり、 ヘビ類系統樹のより基本的系統に位置する。そのため、Hox12遺伝子固有のプライマーバイアスがあった場合はそれを回避できる可能性がある。調査の結果、これら2種のヘビでもHoxC12遺伝子が検出され。従って、ヘビ類ではHoxD12を失ったと考えられる。
カメ
スッポン(Pelodiscus sinensis)は、39のHox遺伝子をもち、トカゲに比べてHoxC3が欠失していた。 これまでに調査された有鱗爬虫類はHoxC3を保持しているので、カメでのHoxC3の欠落がプライマーバイアスによるものかどうかを疑問に感じた。 そこで、別の4種のカメのHox3遺伝子を調査した。黄色の斑点のあるアマゾンカメ(Podocnemis unifilis)、ブタバナカメ(Carettochelys insculpta)、スッポンモドキ(Chrysemys picta)、アカミミガメ(Trachemys scripta) である。これらは、カメの広範な進化をカバーするために特に選択されました。 その結果、HoxC3遺伝子を特定できなかった。これは、カメでのHoxC3の欠失と一致していた。
ワニ
シャムワニ(Crocodylus siamensis)の場合、ほとんどのHox遺伝子は、鶏のHox遺伝子データを参考にして、予想どおり検出された。ただし、いくつかのプライマーの組み合わせを試したが、通常2つの遺伝子(HoxA2とHoxB2)を含むHox2の遺伝子では、1つのHox遺伝子のみが取得された。 HoxA2またはHoxB2の欠失は他の脊椎動物では報告されていないため、HoxA2およびHoxB2にexon1を標的とするプライマーを使用した。その結果、シャムワニについても両方の遺伝子の特定の配列を同定することができた。 さらに、シャムワニではHoxC3は検出されなかった。ワニでのC3の欠失は、中国ワニ(Alligator sinensis:現存するワニの他の主要なクレードの代表)でも確認された。 従って、ワニには39のHox遺伝子があり、それらのHox遺伝子データは哺乳類のそれと同一である。
鳥類
Hoxクラスターのゲノム解析はニワトリ(Gallus gallus)で報告されているが、不完全なゲノム構成により、HoxC4とHoxC5の情報はまだ得られていない。また、ニワトリゲノムのHoxCクラスターの3 '末端にギャップがあるため、ニワトリにHoxC3があるかどうか不明である。さらに、予測されたニワトリHoxC12 のホメオボックスは、他の脊椎動物におけるC12オーソログとは大きく異なり、アセンブリエラーがある可能性があることを見出した。我々のPCR調査では、それぞれのホメオボックス断片を見つけることにより、鶏のHoxC4とHoxC5の存在を確認した。鳥がHoxC3を保持しているかどうかを調査するために、アヒル(Anas platyrhynchos var domestica)とダチョウ(Struthio camelus)のHox3遺伝子を調査した。その結果、HoxC3は検出されず、鳥で欠失していることが示唆された。鶏、アヒル、ダチョウにおけるHox12の再検査により、鳥におけるHoxD12の存在が確認され、他の脊椎動物においてHoxC12とよく保存されているように見えるフラグメントが明らかとなった。従って、鳥類のHoxC4、HoxC5、HoxC12、HoxD12の検出により、鳥類のHox遺伝子データのより完全な情報を提供できた。
哺乳類
有胎盤哺乳動物(例:ヒトとマウス)のHox遺伝子データは明確だが、現存する哺乳動物の他の主要な2群(有袋類、単孔類)の関連情報はまだ報告されていない。従って、ハイイロタンビオポッサム(Monodelphis domestica)とカモノハシ(Ornithorhynchus anatinus)のHox遺伝子のインシリコ調査を行った。両種で、Hox A、B、Dクラスターは、ヒトのクラスターとほぼ同じであった。(HoxB6は、シーケンスのギャップのためにカモノハシで検出されなかった)。 HoxCクラスターの場合、カモノハシにはHoxC5、HoxC4およびその3 '隣接領域をカバーするゲノム配列が欠損している。オポッサムは、HoxC6とHoxC9のフラグメントを除いて、HoxCクラスターのほぼすべてのゲノム配列が欠損していた。 HoxC13-C4の構造がすべての四肢動物で保存されているという観察に基づいて、有袋類と単脚類も同じHoxCメンバーを持っていると推測したくなるが、オポッサムとカモノハシの両方でHoxC4の3 '隣接領域のゲノム配列がまだ欠落しているため、有袋類と単孔類におけるHoxC3の存在/不在に関する結論を導き出すのは時期尚早である。
包括的なシナリオ
我々のデータを他の公開されたデータと組み合わせることで、総鰭類の魚から哺乳類まで、主な肉鰭類系列に沿ってHox遺伝子のデータ変動がどのように発生するかについて、より完全なシナリオを提供できるようになった。 全体として、4クラスターのHox構造はよく保存されている。 始原系統は比較的多くのHox遺伝子メンバーを保持する傾向がある。 HoxA14、B10、C1、C3は、肉鰭類の進化の過程で連続して失われた。 HoxD12とHoxB13は遺伝子欠失の2つのホットスポットであるようで、異なる動物グループがこれらのHox遺伝子を独立して喪失している可能性がある。
Hox, ParaHox, ProtoHox: facts and guesses
J Garcia-Fernàndez、Heredity 94,pages145–152(2005)
要旨
Hox遺伝子クラスターは、世界中の進化および発生生物学者の想像力を魅了した。このレビューでは、始原Hox(protoHox)遺伝子クラスターの複製によるHoxおよび傍Hox (paraHox)遺伝子クラスターの起源と、主要な後生動物の分岐におけるそれらの遺伝子数の変化を批評的にレビューした。既存のデータの再評価と刺胞動物、扁形動物、および脊椎動物の進化の重要な段階における最近の知見は、後生動物の進化の関連事例における起源、構造、およびHox遺伝子数の変化に関する代替となるシナリオの必要性を示唆している。私は反対の見解を議論し(i)始原Hoxクラスターには2つの遺伝子しかなく、一般に信じられている4つではなかったと推論した。当然の結果として、左右相称の起源は新しいHoxおよび傍Hox遺伝子クラスの提案と一致し、そのような分岐理論を促進した。(ii)始原Hoxクラスターの重複は、以前に提案されたように、トランス重複事例(他の染色体からコピーされた)ではなく、シス重複事例(同一染色体内でコピーされた)であり、(iii)祖先の脊椎動物クラスターは14のHox遺伝子を所有しており、13は一般的に想定されていない。これらの仮説は近い将来に検証または反駁される可能性があるが、後生動物におけるHox/傍Hox群の進化についての批判的な議論に役立つと思われる。
背景
ショウジョウバエの2胸郭複合体に関するエドワード・ルイスの独創的な研究(ルイス、1978)以来、Hox遺伝子クラスターは進化論と発生学の生物学者を魅了してきた。Hox遺伝子は、ホメオボックス遺伝子のクラスに属する。これは、発生・分化の多くの側面を調節する転写因子の主要なクラスである(Gellon and McGinnis、1998)。1984年のホメオボックスの発見(McGinnis et al、1984)と同様の遺伝子がハエやマウスと同じくらい多様な動物で同様の方法で作用するという発見は、発生生物学者と進化生物学者の和解を促進した。これは、ダーウィンの「修正を伴う生物分岐:descent with modification」の進化論に触発され、彼の最初の発想(「胚構造の初期のコミュニティは後期のコミュニティを明らかにする」ダーウィン、1859年)に続いた1世紀の苦しみのある発生学と進化論の関係の成果である。ダーウィン以来、実験発生学者は発生学的プロセスの機構を解明することに集中し、進化生物学者は自然集団における遺伝子頻度の変化を追跡してきた。現在、両方の分野の最前線にある進化・発生(エボ・デボ)の新しい分野は、新しい「進化の進化的統合」を求めている(ギルバート、2003)。Hox遺伝子の保存は、エボデボコ研究界に活気を与え、前後関係の体軸をパターン化する保存された遺伝子のセットとして後生動物の「動物型zootype」の概念を定義するのに役立った(Slack et al、1993)。その後すぐに、ボディプランの進化と多様化のために、Hox遺伝子数の変化、配列、および調節の研究が始まった(例:Gellon and McGinnis、1998; Wagner et al、2003; Amores et al、2004)。
Hox遺伝子を発生調節遺伝子の中で特別なものにしたのは、複合する染色体の中での遺伝子の組織化だけでなく、ハエに9つの遺伝子があり、哺乳類では4つのクラスターに39の遺伝子があること、空間的および時間的共直線性の現象であることであった。クラスターの一端にある遺伝子が発現し、胚の前端をパターン化し、クラスターの他端にある遺伝子は後端をパターン化する(Duboule and Dollé、1989)。この空間的共直線性(3‘末端は前方に等しく、5’末端は後方に等しい)は、一部の系統(主に脊椎動物)の時間的共直線性の直接的な結果である。クラスターの3‘末端の遺伝子は、5’末端の遺伝子よりも発生の初期に発現する(Duboule、1994)。したがって、前部から後部に成長する発生系の時間的共線性は、空間的共線性にすぐにつながるということである。共直線性の分子メカニズムはとらえどころのないものであるが、最近は、クロマチンのリモデリングと染色体領域の物理的トポグラフィーが関係しているとされている(Duboule and Deschamps、2004)。
後生動物におけるHox遺伝子の進化はまだ完全には理解されていない。哺乳類と節足動物のHoxクラスターの比較により、後生動物の「上位」の2群である旧口動物と新口動物の最後の共通祖先は、7つの遺伝子で構成される単一のHoxクラスターを持っているという一般的な合意に至った。すなわち、その7つは、前部の2つのパラロガス群(哺乳類のパラロガス群(PG)1–2、ショウジョウバエ口唇側および口唇裂遺伝子)、1つのパラロガス群3(PG3、ショウジョウバエzen遺伝子)、中央部の3つのパラロガス群(PGs4–5、ショウジョウバエ変形および性櫛が減少したもの、 PGs6–8の祖先、ショウジョウバエの触覚脚遺伝子、超胸郭および腹部-A)、および単一の後部のパラログ(PGs9–13の祖先、ショウジョウバエの腹部-B)。前口(旧口)と後口(新口)の両方の系統では、この元のクラスターは明確な進化の経路をたどり、現在の複合体の構成を説明する中央領域と後方領域に追加の遺伝子のタンデム重複がある(de Rosa et al、1999; Ferrier andMinguillón、 2003)。
1998年に傍(para)Hoxクラスター(Brooke et al、1998)が発見されるまでは、Hox遺伝子はHoxクラスターの進化的相同複合体であった。傍Hox遺伝子の発見は、4つの遺伝子の仮定の始原Hoxクラスターが動物の進化の初期に複製され、2つの双子クラスターを生じさせることを示していた。これらは、ショウジョウバエの8つの遺伝子、または哺乳類の13のパラロガス群に拡張された始原Hoxクラスターと、1つのメンバーを失い、少なくとも3つの遺伝子複合体が頭索動物と脊椎動物におけるシス重複によって発生したことである。ここでは、Hoxと傍Hox遺伝子の最近のデータと異なる系統のクラスター内容をまとめ、Hoxと傍Hox遺伝子クラスターの進化に関するさまざまな仮説について説明する。特に、(i)左右相称動物とカンブリア紀生物爆発の起源に伴う進化の変化、Hoxと傍Hoxクラスターの起源と元の構造、(ii)Hoxと傍Hoxの「付随する」遺伝子クラスター、または始原Hoxクラスターの複製がシスまたはトランス事例であったかどうか、および(iii)脊椎動物の系統におけるHox遺伝子クラスターの基本的な内容についてである。
仮説1 2倍体か4倍体か?始原Hoxクラスター
両生類のゲノム解析から、3つのHox遺伝子に密接に関連する遺伝子AmphiGsh、AmphiXloxおよびAmphiCdxが発見された。それらの遺伝子のHox遺伝子クラスターとの系統関係から、始原Hoxクラスター仮説へとつながっていった(Brooke et al、1998)。塩基配列の類似性に基づいて、Hox遺伝子は4つのグループに分類できる。前部パラログ群(PGs1〜2)、パラログ群3、中央パラログ群(PGs4〜8)および後部パラログ群(PGs9〜13)である。Gsh遺伝子は他の傍Hox遺伝子よりも前部パラログ群Hox遺伝子に密接に関連している。XloxはHoxPGs3に、CdxはHox後部パラログ群の遺伝子に類似している。従って、モデルは4つの遺伝子を持つ始原Hox複合体を予測させる:前部では、前部Hox遺伝子の祖先(PG1 / 2)および前部傍Hox(Gsh)遺伝子、始原Hox遺伝子PG3とXlox)、中央の始原Hox遺伝子(中央のHox遺伝子(PG4/8)の祖先、中央の傍Hox遺伝子は始原Hoxクラスター複製の直後に失われた)、および後部の始原Hox遺伝子(後方のHox遺伝子PG9 / 13の祖先)およびCdx)。傍Hoxクラスターに関する新しい発見(例:Finnerty and Martindale、1999; Yanze et al、2001; Ferrier and Holland、2003; Cook et al、2004)およびHox / 傍Hoxの進化に関するレビュー(例:Ferrier and Holland、2001;Martínez and Amemiya) 、2002; Ferrier andMinguillón、2003)は常に、4つの遺伝子を持つ始原Hoxクラスターを持つ進化的フレームワークを提示している。ここでは、始原Hoxクラスターとして、原始的なHoxクラスターと傍Hoxクラスター構造を代替シナリオとして提案する。これは、利用可能なデータの再検討と下位後生動物に関する最近の発見を考慮したためである。
Brooke et al(1998)は、主要な前口動物および後口動物門が出現したカンブリア紀の生物爆発と同時に、始原Hoxクラスターが複製されたことを提案した。しかし、すぐに二胚葉の刺胞動物が既にHox遺伝子と傍Hox遺伝子を持っていることが明らかとなった(Finnerty and Martindale、1999)。したがって、始原Hoxクラスターの重複は、刺胞動物と三胚葉動物の分岐前に発生している必要がある。二胚葉動物におけるHoxと傍Hox遺伝子の検索は非常に熱心に行われた。刺胞動物門、花虫綱、水生動物、箱虫綱(ハコクラゲ綱)、および鉢虫綱の異なる種が、Hoxおよび傍Hox遺伝子について広範囲にわたって検索された。その結果、主要なコンセンサスが生まれつつある(Finnerty、2003年に要約; Finnerty et al、2004):刺胞動物のHox/傍Hoxクラスターの基本的な内容は、1つの前部と1つの後部Hox遺伝子、及び1つの前部と1つの後部傍Hox遺伝子からなる。激しい努力にもかかわらず、PG3遺伝子や中央のHox遺伝子や傍Hox遺伝子は、刺胞動物種で発見されなかった。したがって、2つの結論が可能である:(i)4遺伝子の始原Hoxクラスターモデルによると、PG3Hoxと傍Hoxはすべての刺胞動物で失われ、中央の傍Hoxはすべての刺胞動物と左右相称動物で失われた、または(ii) 始原Hoxクラスターと原始的なHoxと傍Hoxクラスターには、前方遺伝子と後方遺伝子しかなかった。つまり、始原Hoxクラスターは、2つの遺伝子だけで構成されていた。
4遺伝子モデルは、8億年以上前に分岐したホメオドメイン蛋白質の60アミノ酸のみ(またはホメオドメインと5つの隣接アミノ酸)の系統学的分析によってサポートされている(Banerjee-BasuおよびBaxevanis 2001; Peterson et al 2004 )。Hox 遺伝子に対する傍Hox遺伝子の特定の群のブートストラップ値は、40〜70%の範囲である(Brooke et al、1998; Finnerty and Martindale、1999; Banerjee-Basu and Baxevanis、2001;MinguillónandGarcia-Fernàndez、2003)。これらの値は、初期の分岐クレード間の関係を確立するための18SリボゾームRNAまたは完全蛋白質配列を用いた系統解析の信頼率をはるかに下回っている(Hillis and Bull、1993)。さらに、4遺伝子クラスターモデルでは必然的に、2つのクラスのHox遺伝子(PGs3と中央)および1つ(PGs3)または2つ(PGs3と中央)の傍Hox遺伝子が独立して刺胞動物系列で失われたことを意味する。さらに、刺胞動物では4つの遺伝子ではなく2つの遺伝子が進化的考察とより一致している可能性がある。左右相称動物のHox遺伝子は、前後の体軸をパターン化している。他方、刺胞動物のHox遺伝子は、口部-反口部を貫く体軸に沿って互い違いの発現を示すが(Finnerty、2003; Finnerty et al、2004)、そのような発現の極性に関するデータは(4遺伝子クラスターモデルでは)不可解である(Masuda-Nakagawa et al、2000; Yanze et al、2001; Finnerty 、2003; Finnerty et al、2004)。結果的に刺胞動物におけるHox遺伝子の共線性は、もしあったとしても、2つのHox遺伝子だけで事実と調和させることは困難である。
初期の左右相称動物のHox補完遺伝子に関する最近のデータは、代替シナリオを想定するのに役立つ。分子系統学における最近の進歩により、以前、前口動物の扁形動物門(Platyhelminthes)のメンバーであった無腸動物(扁形動物プラナリアに近縁の皮中神経目Acoela Nemertodermatida)が、最も古い現存する左右相称動物クレードに属することが明らかにされた(Ruiz-Trillo et al、1999、Baguñaand Riutort、2004でレビュー)。したがって、無腸動物Acoelomorphaの特別な中間の位置(刺胞動物と左右相称動物の間)は、単純な、しかし分断化されていない、新しい左右相称動物として、Hoxクラスターの進化を理解するのに有用である。無腸動物(Acoels)には、「標準的で」より高度な左右相称動物の持つHoxおよび傍Hoxクラスター(7つのHox遺伝子、3つの傍Hox遺伝子)が存在するか?それとも初期バージョンのHoxおよび傍Hoxクラスターしか持っていないか?刺胞動物/左右相称動物への移行は、前後の体軸の多様化に関与しているため、Hoxおよび傍Hox遺伝子の数の増加を伴っている可能性が考えられる。4遺伝子モデルが正しく、刺胞動物と左右相称動物の共通の祖先にすでに4つのHox遺伝子と3つから4つの傍Hox遺伝子があった場合、無腸動物でより複雑なクラスターを見つけることが期待される(たとえば前口動物と後口動物の最後の共通の祖先が、最大7つの遺伝子のHoxクラスター持つ)。他方、2遺伝子モデルが正しく、刺胞動物がこの古い遺伝子発現状態を示す場合、無腸動物クラスターでは3つ以上の遺伝子が見つかると予想される。非常に最近のデータ(Cook et al、2004; Baguñàand Riutort、2004)は、無腸動物が4つの遺伝子(1つは前方、1つはPGs3、1つは中央、1つは後方)のHoxクラスターと傍Hoxクラスターの3つの遺伝子を持っていることを強く示唆している。したがって、4遺伝子モデルの下では、各標準クラスの1つの動物は、刺胞動物/左右相称動物への移行に関してHoxまたは傍Hox補完遺伝子の増加を伴わなかったことになる。しかし、2遺伝子モデルが正しい場合、刺胞動物/左右相称動物への移行は、2つのHox遺伝子(PGs3および中央)、および1つの傍Hox遺伝子(Xlox)の出現を伴っていた。少なくとも3つの遺伝子を持つクラスターが、刺胞動物の初期の2遺伝子クラスターとは対照的に、新しく獲得した左右相称の前後軸を異なるパターンでパターン化する強力なツールだったと想像することは、魅力的な考えである
中間の仮説、たとえば3遺伝子(前部、PDs3、後部)を含む始原Hoxクラスター説を先に進めることも可能である。 この場合、刺胞動物ではPG3とXloxを失い、左右相称動物の出現に伴う遺伝子の複雑さの増加は、中央 Hox遺伝子のみに起こったことと関連する。 このモデルは、PG3とXloxの密接な系統関係により、より良く適合する。 ただし、まだ2つの遺伝子(PG3とXlox)が刺胞動物の系統で独立して失われたこと説明する必要がある。 2遺伝子モデルでは、必要な唯一の前提は、新しい遺伝子の発現がボディプランの複雑さの増加に関連していること、およびXlox / PG3の系統分類が人為的なものであるか、説明の難しい機能的収束によるものであるかということである。
要約すると(i)刺胞動物に対する集中的な検索により、彼らが2つの遺伝子からなるHoxおよび傍Hoxクラスターを持つこと、(ii)初期の左右相称動物がそれぞれ4つおよび3つの遺伝子を持つHoxおよび傍Hoxクラスターを持っているという発見、(iii)基礎をなす複雑な左右相称動物の持つ拡張されたHoxクラスター(7つのHox遺伝子)、(iv)Hoxおよび傍Hoxグループの低いブートストラップ値、および(v)Hoxおよび傍Hox遺伝子の数はボディプランの複雑さと前後の多様化の増加と相関している可能性が高いという示唆が得られている。
私は主要な後生動物の進化的な移行のいくつかについて次の仮説を提案する。最初に、2つの遺伝子を持つ始原Hoxクラスターは、刺胞動物/左右相称動物への移行に先立って複製された。現在の刺胞動物には、2-Hox、2-傍Hoxの始原クラスターの直接の子孫であるクラスターが存在する。第2に、刺胞動物/両生類の移行には、Hoxクラスターと傍Hoxクラスターの独立したタンデム複製による拡張が伴った:Hoxクラスターでは、PG3と中央グループの遺伝子は、前部遺伝子の複製によって、またはシャッフルと多様化によって発生した; 傍Hoxクラスターでは、Xloxは後部の傍Hox遺伝子のタンデム複製によってできた。第3に、左右相称動物の進化の初期段階で、Hox複合体はクラスターの中央部分でさらに遺伝子重複を受け、PG4、PG5、およびPGs6–8のファウンダー遺伝子を産生した。前口動物および後口動物系統では、それぞれのHoxクラスターの内容を複製することにより、中央部および後部のHox遺伝子がまだ生成されている。 このフレームワークは、後生動物の主要な移行時におけるHox / 傍Hox遺伝子の内容の変更を直接的に呼び起こす。(i)刺胞動物 / 左右相称動物の分岐前の始原Hoxクラスターの複製、(ii)Hoxと傍Hoxクラスターの拡大は、左右相称動物の起源と一致して起きている。(iii)複雑な左右相称動物の出現と一致するHoxクラスターのさらなる中央部遺伝子拡大。 遺伝子数のこれらの変化がこれらの主要な移行の原因であったと推測するのは魅力的な考えである。
ゲノムが2つまたは4つのHox遺伝子を含んでいるどうかに関係なく、始原Hoxクラスターは刺胞動物から左右相称動物への分岐よりも早く複製されたことは間違いない。始原Hoxクラスターがどこで作成または複製されたかは不明である。下部後生動物、すなわち有節動物、平板動物、およびカイメンの正しい分岐は明確ではない(例:Martindale et al、2002; Ender and Schierwater、2003)、しかし、これらの動物では始原Hoxクラスターの検索には成功していない。 単一のHox様遺伝子が有節動物(Ctenophores、クシクラゲ(Finnerty et al、1996))に存在する可能性があるが、完全なHox様遺伝子はカイメンでは報告されていない。 平板動物の遺伝子Trox2が起源のHox / 傍Hox遺伝子(UrProtoHox遺伝子)(Jakob et al、2004)に由来する可能性があるという最近の主張は、刺激的ではあるが、さらなる検索が必要である。
推論2:Hox遺伝子はシス/トランス重複か?Hox,傍Hoxクラスターのタンデム重複と結合アレイは?
Hox様遺伝子Evxは、脊椎動物(Dush and Martin、1992; Amores et al、1998; Powers and Amemiya、2004)および刺胞動物(Miller and Miles、1993)においてHoxクラスターと密接に関連している。EvxとHox遺伝子の連鎖を説明するための最も説得力のある仮説は、刺胞動物と左右相称動物の分岐前の始原Hoxクラスターでは、すでにEvxと密接に関連していたということである。Pollard and Holland(2000)によるヒトゲノムの分析は、ショウジョウバエの触覚脚ファミリーの4つのHoxクラスター、拡張されたHoxクラスター(Hoxクラスター及び関連するHox遺伝子EvxとMox)、NKLクラスター(これには次のようなHox遺伝子が含まれる: Nkx、Msx、Dlx、Tlx Emx、またはLbx)、およびEHGboxクラスター(En、HB9、Gbxを含む)は、タンデム遺伝子重複および後生動物進化の初期の祖先のUrArcheHox遺伝子からのクラスター重複によって発生した。さらに、そのようなクラスターの証拠は、ナメクジウオとショウジョウバエのゲノムにも見られる(Castro and Holland、2003; Luke et al、2003)。そのようなモデルでは、始原Hox、EHGbox、およびNKLクラスターは、各クラスのファウンダー遺伝子(始原Hox、始原EHGbox、および始原NKL遺伝子)の祖先クラスターからの連続したタンデム複製によって生じたと思われる。したがって、始原Hoxクラスター(トランス重複)は非タンデムに複製され、始原HoxクラスターはEHGboxおよびNKLの原始クラスターの隣に残るが、原始の傍Hoxクラスターはゲノムの他の位置にジャンプする。この提案は、EvxとMoxがHoxクラスターの同じ端(5 '端)にマッピングされた2000年のヒトゲノムマッピングに基づいており、EvxとMoxの両方がHoxクラスターの遺伝子のタンデム複製から生じたと仮定した。
繰り返しになるが、系統学的分析は不可解であった:Evx遺伝子はHox/傍Hox遺伝子クレードの基礎となる(Gauchat et al、2000; Kourakis and Martindale、2000)。この位置は、Hov/傍Hox始原クラスターを生成する複製イベントの前にEvxが出現したことを示唆している。Moxクラスは系統解析にほとんど含まれておらず、不明な傍Hox中心遺伝子というように漠然と呼ばれている(Gauchat et al、2000; Hill et al、2003)。2つの広範な系統学的分析(Banerjee-BasuおよびBaxevanis、2001;MinguillónおよびGarcia-Fernàndez、2003)は、EvxとMoxが密接に関連しており、Hox/傍Hoxグループのクレードの基礎を形成していることを示唆している。したがって、EvxとMoxは、Hox/傍Hoxクラスターの生成の複製イベントに戻る祖先(Evx/Mox祖先または始原MoEve)の複製によって発生した。したがって、系統発生データは、始原MoEve遺伝子が始原Hoxクラスターに隣接していたことを示唆している。
現在の系統発生データに適合する始原MoEve/始原Hoxクラスター(ここではHoxのようなクラスターと呼ばれている)の特定の複製事例を追跡するために、いくつかのシナリオが想定されている。(i)始原MoEveのEvxとMoxへの初期のシス重複、および後で、PollardとHolland(2000)によって提案された始原Hoxクラスターのトランス重複。これにより、EvxとMoxの両方がHoxクラスターの同じ側の隣に配置された。(ii)Hoxのようなクラスターの単一のトランス重複事象の結果としてのEvx、Mox、Hox、および傍Hoxの形成。これにより、次にEvxがHoxクラスターの5 '端に残り、Moxが傍Hoxクラスターの5'端に残る。現在まで、Moxはどの系統でも傍Hoxクラスターの横には検出されなかった。(iii)拡張されたHoxのようなクラスターの単一のシス重複事象の結果としてのEvx、Mox、Hox、および傍Hoxの形成。これは、Evx-Hox-Mox-傍Hoxの配列を持つ「結合されたHoxのようなクラスター」になる(EvxはHoxクラスターの5 '端にあり、MoxはHoxクラスターの3'端にあることに注意。および傍Hoxクラスターの5 '末端)。繰り返すが、そのような遺伝子の完全な配列はどの系統にも見られない。しかし、ヒトとマウスのゲノムの明確なマッピング(Spring、2002)は、Moxが信じられていたのとは逆に、Hoxクラスターの3 '末端に約5Mbあることを示した。この場所は、傍Hoxクラスターのいずれかの側で染色体の破損をモデルに含めるだけで、結合されたHoxのようなクラスターのタンデム重複事象の仮説と一致する。そのような破損は、EvxとMoxをHoxクラスターの両側に残したまま、傍Hoxクラスターを分離した形になる。タンデム重複とそれに続く染色体切断により、系統発生解析および現在の連鎖解析がうまく二乗される。興味深いことに、尾索動物の ワカレオタマボヤ(Oikopleura dioca)のHox内容の特徴付けは、CdxがHox1に連鎖していることを示している(Seo et al、2004)。この連鎖が祖先の状態の残骸なのか、それとも単なる偶然の幸運なのか、またHox/傍Hoxの破損が進化の初期に一度だけ発生したのか、それとも何度も発生したのかは不明。
タンデム重複とそれに続く始原Hoxのようなクラスターの染色体切断により、一緒にリンクされている3つのHoxクラスター(始原Hox、EHGbox、NKL)のモデルに改良が加えられる。これは、後生動物の進化の初期(確かに刺胞動物/左右相称動物の分岐の前)、4つのHoxクラスター(EGHbox、Hox-with Evx-、傍Hox-with Mox-、およびNKL)の拡張配列、4触覚脚遺伝子のような始原Hoxクラスターの配列が存在している。 重複重複イベントがなかったことは、重複がAntp(触覚脚遺伝子)のようなHoxクラスターを生成することは、ゲノムの倍数化の産物ではなかったことを示している。拡大によるものであり、刺胞動物/左右相称動物の分岐に先立つ完全なゲノム重複はありそうにない。
推論3:13あるいは14遺伝子、脊椎動物はHox14遺伝子を持つか?
哺乳類には4つのHoxクラスターがあり、単一のクラスターから脊椎動物の進化の初期に重複した後の遺伝子損失の結果として、PG1からPG13までの13のパラロガスグループ(PG)の代表が存在する。それゆえ、基盤となる脊椎動物クラスターは、各PGの1つである13個のHox遺伝子を保有しているはずであると考えられていた。脊椎動物の無脊椎動物姉妹グループである原索動物ナメクジウオ(amphioxus)の単一のHoxクラスターの最初の発見(Garcia-Fernàndezand Holland、1994)は、モデルにうまく適合した。しかし、頭索動物クラスターのさらなる分析により、驚くべきことに、13遺伝子クラスターではなく、14番目の遺伝子AmphiHox-14が明らかになった(Ferrier et al、2000)。脊索動物の後部Hox遺伝子(PG9–PG13または14)の系統学的分析は、それらのより高い進化率、後部柔軟性と呼ばれる現象(Ferrier et al、2000)によって不明瞭になっている。そのような分析では、AmphiHox-14がナメクジウオゲノムの系統固有の重複を表すのか、脊椎動物のパラロガスグループの残骸であったのか、その後の脊椎動物の進化の初期に失われたのか明らかになっていない。
ごく最近、Powers and Amemiya(2004)による発見により、ナメクジウオのデータが強化された。シーラカンスのHoxAクラスターとツノザメのHoxDクラスターの分析により、それぞれの動物種に追加の14番目の遺伝子A14とD14が明らかになった。さらに、ツノザメのHoxAクラスターにもHoxa14遺伝子の偽遺伝子が見つかった。サメは初期の顎口類の代表であり、シーラカンスは初期の肉鰭類(総鰭類の魚で、哺乳類を含む四肢動物)である。これらの発見は必然的に、顎口類(魚類)の共通の祖先がすでに14番目のHox遺伝子を持っていたことを意味する。これらの新しい14番目の遺伝子の系統学的分析では、おそらくHox遺伝子の後方柔軟性の別の結果として、AmphiHox-14が脊椎動物の14番目の遺伝子の前オーソログであるかどうかを解決できない。ただし、クラスター内での位置とイントロンの共有(Ferrier et al、2000; Powers and Amemiya、2004)は、単一の祖先の脊椎動物、脊索動物、または後口動物のクラスターに14PGが含まれているという仮説を支持するものである。
Hox14遺伝子モデルは、下部脊椎動物(無顎類:Agnathans)、下部脊索動物(尾索動物:Urochordates)、または他の後口動物(半脊索動物および棘皮動物)でこのような遺伝子を検索することによって調査できる。現在の利用可能なデータでは問題の解決に役立たない。 無顎類、尾索動物、棘皮動物および半索動物(Hemichordates)に関する公開された利用可能なデータは、複数の後部遺伝子PGs9〜13を示唆している。これは、無顎類で少なくとも5例(Fried et al、2003)、尾索類で最大6例(Spagnuolo et al、2003; Seo et al、2004)、棘皮動物で少なくとも4例(Martínezet al、1999)、半索動物で少なくとも3例(Peterson、2004)の検索結果である。後部Hox遺伝子の柔軟性は、再び、後口動物の後部遺伝子間の特定の関係の解決の妨げとなっている。要約すると、後口動物には複数の後方遺伝子があり、一部の系統では最大6つのメンバー(PGs9–14)があり、現在のデータでは、脊椎動物のゲノムで2回複製された元のクラスターには、最も単純な仮説として14の遺伝子があったことが示されている。このようなクラスターの構成は、頭索類のナメクジウオの単一のクラスターと完全に一致する。
結論
最近のデータの分析により、後生動物の進化の重要な交差点にいて、よく知られているHoxと傍Hoxクラスターの起源と構造に関する新しい仮説が導かれる。これらの仮説は、広範な関連する情報によって補足された系統発生情報に基づいており、最も節約されたモデルである。以下は、私の提案の要約である。
i)基本の始原Hoxクラスターには、前方遺伝子と後方遺伝子の2つの遺伝子しか存在しない。 それらは、刺胞動物/左右相称動物の分岐以前に重複した。対称性の起源、左右相称動物の起源、カンブリア紀の生物爆発のような主要な後生動物の移行は、始原Hox、Hox、または傍Hoxクラスターの複雑さの増加に伴って起こった。
ii)4つのHoxクラスター(EgHbox、拡張Hox、傍Hox、NKL)の複雑な配列が後生動物の進化の初期に存在した。
iii)祖先の脊椎動物クラスターは14のパラロガス群(PG)に拡大していた。
これらの仮説は、それらの後生動物の移行を明らかにする動物のゲノムの広範な分析を必要とするが、検証されるべきである。
2019年春①の試験問題回答です
正しい文章には〇を、間違っている文章は、下線部を正しい言葉に置き換えなさい。
正しい文章は1つだけです。2つ以上〇をつけると減点になります。
〇を2つ以上付けた学生さんがいます。国家試験では本当に0点になります。注意してください。
第1回講義から
1、地球上に最初に出現した細菌は化学合成独立栄養細菌と考えられている。( 〇 )
2、好塩菌やメタン菌、好熱菌を含む古細菌は、真菌から38億年くらい前に分岐したと考えられる。(真正細菌)
3、光合成を行う細菌の中で酸素を産生する細菌はヘリオバクテリア、化石はストロマトライトとして残っている。(シアノバクテリア)
4、フレミングが見つけたカナマイシンはブドウ球菌を培養しているときに飛び込んだアオカビが産生した。( ペニシリン )
5、大腸菌の雌雄を決めるのはP因子であり性線毛を産生するプラスミドである。( F因子 )
第2回講義から
1、真核生物が原核生物と違うのは核膜や、ミトコンドリア、葉緑体、リボソーム等の細胞内小器官を持つ
ことである。(リボゾーム削除
2、一般に原核生物と真核生物の共生を一次共生、真核生物同士の共生を二次共生という。( 〇 )
3、原生動物の分岐は根足虫、鞭毛虫、胞子虫(アピコンプレックス群)、そして繊毛虫の順である。(鞭
毛、根足、胞子、繊毛の順)
4、トリパノソーマ、赤痢アメーバ、マラリア原虫は無性生殖のみだが、ゾウリムシは有性生殖する。(マラ
リア原虫は有性生殖 )
5、多細胞動物のもとになった単細胞性の原核生物としてはゲノム解析の結果、立襟鞭毛虫が考えられ
ている。( 真核生物 )
第3回講義から
1、真菌類の中で最も多くの多様性を持つのが子嚢菌門、次いで担子菌門、最も少ないのがツボカビ門
である。(接合菌門)
2、1胚葉の側生動物門は海綿のみ、2胚葉の刺胞動物にはクラゲ、イソギンチャク、ヒトデなどがある。( ヒトデ削除 )
3、ヒドラは、表皮筋細胞、刺胞細胞、間細胞、消化細胞、腺細胞を持つが神経細胞はない。(ある )
4、プラナリアは3胚葉性の扁形動物で無体腔動物であるが、目を持っている。( 〇 )
5、自由生活のプラナリアは扁形動物であるが、寄生性の糸状虫(回虫)、吸虫(肝蛭)、条虫(サナダムシ)は線形動物である。(吸虫、条虫削除)
第4回講義から
1、生物爆発といわれるカンブリア紀(5.4~4.9億年)以前の動物はオルドビス動物群に代表される薄い体
の生き物である。(エディアカラ)
2、扁形動物から脱皮動物、甲殻類、昆虫へと分岐した。昆虫の一部は陸生となり、空を飛ぶものが出現した。( 線形動物 )
3、エビやカニは自然免疫しか持たないが、ヘモシアニンを持つ。蜜蜂は血色素を持たないが、特異免疫を持つ。(特異免疫も持たない)
4、昆虫は解放血管系を持つ。腎臓に匹敵する排出器官はマルピーギ管で腸に開口する。(〇 )
5、蜜蜂では1匹の女王バチ、数万の雄の防衛バチと数万の雌の働きバチで社会を構成する。(数百~数千、交尾用)
第5回講義から
1、ホメオボックス遺伝子から見ると頭索動物のホヤが基本となり、脊椎動物では、遺伝子は4倍になっている。(ナメクジウオ )
2、脊椎動物に属する無顎類のヌタウナギは大きな嗅球を持つが中脳を欠いている。(小脳)
3、硬骨魚類には淡水魚と海水魚がいるが、外界との浸透圧調節に働くのが尿素細胞である。(塩類細
胞)
4、魚類から両生類となり、陸に上がり、浮力の消失・鰓呼吸から肺と皮膚呼吸などに対応することとなった。( 〇 )
5、爬虫類の多くは、鳥類と同様に尿を水に不溶な尿素としてクロアカ(総排泄口)から排出する。(尿酸)
第6回講義から
1、異なる種の染色体間で、遺伝子が同じ順番で配置されていること、もしくはその領域をオーソログという。(シンテニー )
2、内温性(恒温性)動物の短所は、体温を安定させるためには産熱と冷却を行わねばならない。(〇 )
3、鳥類の免疫中枢は哺乳類と異なり、T細胞は骨髄、B細胞はファブリキウス嚢で分化、成熟する。( T細胞は胸腺 )
4、オオコウモリは果食性で、立体視、エコロケーションをおこない、昼行性種がいる点でココウモリと違う。(エコロケーション削除)
5、古細菌や真正細菌、原生生物などは微生物型ロドプシンと呼ばれる圧力受容型膜蛋白を持っている。( 光受容型 )
第7回講義から
1、250万年前に氷河期に入り、草原は乾燥し石器を使用する群が出現、草食から肉食へ、猿人から原
人へと分岐した。( 〇 )
2、ヒトは、大人になっても、姿や行動が類人猿の子供によく似ており、幼形成熟(パラログ)といえる。(
ネオテニー )
3、ヒトの脳は体の約2%の重量であるが、ブドウ糖消費量は15~20%、酸素消費量は10%である。( 20% )
4、ヒトは発語に関連するブローカ言語野と視覚情報処理のウェルニッケ言語野が発達した。( 聴覚 )
5、ヒトの大脳皮質で拡大した側頭葉は自意識、意志、推測、やる気、戦略などの機能を果たしていると考えられている。(前頭葉 )
おまけ。面白かった質問から
1、鳥類が爬虫類から分岐したのは、ほ乳類が爬虫類から分岐したよりもかなり早いと考えられます。
(遅い)
2、マラリア原虫、トリパノソーマ等の腸管寄生原虫は、体表の細胞膜にある輸送蛋白質で宿主から栄養
素を獲得する。( 血液 )
3、葉緑体の外膜は細胞由来、内膜は細菌由来である。( 細菌由来 )
4、蜜蜂では32本の染色体をもつのが雌(女王バチ、働きバチ)、16本の染色体をもつのが雄バチです。(〇)
5、ネアンデルタール人の遺伝子はヨーロッパ人やアジア人には入っているがアメリカ人には入っていない。( アフリカ人 )
妻と作った人形。
娘の修学旅行の写真をもとにしました。
妻と作った人形。
娘の修学旅行の写真をもとにしました。

オリジナルの写真です

娘のドイツ時代のカーニバルの写真です。大家さんは子ネズミちゃん「モイスヒェン」といっていました。
下の人形は妻の作品です。


先日、妻の作品が創刊700号記念家庭画報大賞の佳作に入りました。
題「何して遊ぼう」です。


妻が、稽古に通い、粘土で作った作品です。昨年、東京フォーラムで、他の生徒さんと一緒に展示されました、「仙人草」
(水やり不要です)。

妻の人形作品です。
ドイツ時代の香代の幼稚園の友達です

ある夏のスナップです。妻の父母、娘、甥たちの集合写真から作りました。
