 吉川泰弘の
ホームページへようこそ!
吉川泰弘の
ホームページへようこそ!
2021年3月4日「進化免疫学」が無事に15回の講義をすべてVODにし、You Tubeに載せました。一応完成したので、前の項目に移動しました。
病原体の科学が病原体から見た世界、生命史という視点であったのに対し、生体防御学は免疫学を基礎に置いた宿主の側の対応を進化論的視点から論じるもので、その意味では「進化と免疫学」とでもいうものです。系統発生が理解できれば、免疫反応の理解は比較的簡単です。神経系やゲノムと同様に「過去のシステムを消さないで、進化途上の体制を残したまま上に積み上げた系が免疫系です」。免疫系の理解にはこのキーワードが非常に大切です。概論のあと、免疫の特性、感染症と免疫の歴史をたどりながら総論を終わります。
各論では自然免疫、獲得免疫(細胞性免疫、液性免疫、ネットワーク)を講義します。生命科学における免疫現象の生化学、分子生物学、遺伝学、ゲノムという、最も面白い分野です。最後に自己免疫病、アレルギーといった免疫現象の負の側面、遺伝性免疫不全と感染による免疫不全症で講義は終わります。
ホームページを開く機会に、この講義スライドをアップしました。スライドを見ると5年前になっていますが、今でもかなりの人が、ホームページの「進化免疫学」を見てくれています。コロナウイルスの影響でVODを作ることを覚えたので、「進化免疫学」講義としてVODを残そうと思いました。2020年秋学期の講義のVOD作成の合間を縫ってYou Tubeにのせておきます。
第1回は免疫の概論です。生体防御の基盤となる自然免疫、特異免疫を紹介します。
1の1イントロ:https://www.youtube.com/watch?v=Z4Gdqele2rw
1の2自然免疫:https://www.youtube.com/watch?v=sX684gKHBxI&t=5s
1の3特異免疫:https://www.youtube.com/watch?v=n2zWTl20DUU&t=314s
2回は、免疫の特性である免疫記憶、自己認識、特異性、クローン、ネットワークの紹介
2の1免疫記憶:https://www.youtube.com/watch?v=MICgRBMreAY
2の2免疫寛容:https://www.youtube.com/watch?v=qNubM0tQf7M
2の3ネットワーク:https://www.youtube.com/watch?v=ZF7Bih-lRKQ
3回は生体防御に入る前、免疫概念につながる人類の感染症の歴史を振り返ります
3の1感染症史:https://www.youtube.com/watch?v=7ad_zX_hTz4
3の2顕微鏡:https://www.youtube.com/watch?v=Jbq_MluFyhg&t=51s
3の3天然痘:https://www.youtube.com/watch?v=z6E4xBrHjWc&t=17s
4回はパスツールが初めて、免疫による防御:ワクチンを開発していく過程のエピソード
4の1家禽コレラ:https://www.youtube.com/watch?v=M9eLt2dfvSU&t=23s
4の2炭疽:https://www.youtube.com/watch?v=rt40-MCQ16g&t=1022s
4の3狂犬病:https://www.youtube.com/watch?v=g0VjSOSZZRE
5回は消毒から微生物の不活化、抗毒素血清の作成から抗体のイメージへの発展過程です
5の1消毒:https://www.youtube.com/watch?v=ZNRcvlVcQlg
5の2破傷風:https://www.youtube.com/watch?v=23pcvXwk1YM
5の3ジフテリア:https://www.youtube.com/watch?v=6EXiEFBKVFA&t=400s
5の4ワクチン:https://www.youtube.com/watch?v=mytGl6WcdSo&t=4s
6回は20世紀の分子生物学をリードした免疫学をノーベル賞の受賞研究から紹介します
6の1ノーベル賞:https://www.youtube.com/watch?v=SJ3SWfXvVFE&t=173s
6の2分子前期:https://www.youtube.com/watch?v=6fgbkFh1O9I&t=826s
6の3分子後期:https://www.youtube.com/watch?v=MJA_88h42Wo&t=432s
7回からは進化免疫学の各論です。自然免疫の前半、レクチンと補体経路、両者の関係
7の1レクチン:https://www.youtube.com/watch?v=Oel4mFRNMU0&t=809s
7の2補体:https://www.youtube.com/watch?v=gghNom1Fiak&t=1052s
8回は自然免疫の後半、抗菌ペプチド、トール様受容体、インターフェロンです
8の1ペプチド:https://www.youtube.com/watch?v=K-j9v0JeYfQ&t=443s
8の2TLR:https://www.youtube.com/watch?v=KEdw74Ni6kQ&t=415s
8の3IFN:https://www.youtube.com/watch?v=-Yt_RNjmkB4&t=14s
9回は細胞性免疫への道を説明します。これまで魚類でしたが特異免疫の原型は円口類
9の1振返り:https://www.youtube.com/watch?v=ITX217aAX44
9の2無顎類:https://www.youtube.com/watch?v=4bsFZoMUlgc&t=3s
9の3NKNKT:https://www.youtube.com/watch?v=fVe7_TEaiIo
9の4細胞性免疫:https://www.youtube.com/watch?v=D-VJKMIgzds
10回は液性免疫のイントロ、抗体構造、移行抗体、ゲノムの再構成について説明します
10の1液性免疫概論:https://www.youtube.com/watch?v=-ghNDCcYZFE
10の2抗体構造:https://www.youtube.com/watch?v=X4rdd0wws4g
10の3移行抗体:https://www.youtube.com/watch?v=Me3dA9_M4K0
10の4ゲノム再構成:https://www.youtube.com/watch?v=9zxIrQwGWy8
11回は、免疫ネットワークの仕組みです。サイトカイン,H2, HLA, MHCの紹介
11の1サイトカイン:https://www.youtube.com/watch?v=PfO0myeV-6I
11の2H2抗原:https://www.youtube.com/watch?v=4lwF0GLkRYY
11の3HLA:https://www.youtube.com/watch?v=cAvMtNLYtp4
11の4MHC: https://www.youtube.com/watch?v=eBv89oB4zOM&t=0s
12回は自己免疫病です。自己免疫病の機序、全身性・臓器別自己免疫病の紹介
12の1自己免疫病:https://www.youtube.com/watch?v=MUWtSmIfZvw
12の2全身性AID:https://www.youtube.com/watch?v=dtDswYGfeJs
12の3臓器別AID: https://www.youtube.com/watch?v=ajkBcidayTg
13回はアレルギー特集。アナフィラキシー、アトピー、I型~IV型、化学物質、免疫検査法
13 の1アレルギー概論:https://www.youtube.com/watch?v=6FxKboDGhvo
13の2化学物質:https://www.youtube.com/watch?v=c-2mWpb9YMs&t=0s
13の3免疫毒性試験:https://www.youtube.com/watch?v=DckVzoejRrQ&t=0s
14回は遺伝性免疫不全症のレビュー。イントロ、自然免疫不全症、特異免疫不全症。
14の1イントロ:https://www.youtube.com/watch?v=W9jmJi-sOqU
14の2自然免疫:https://www.youtube.com/watch?v=5yGvYWnUaqY
14の3特異免疫:https://www.youtube.com/watch?v=NtCcXKGd_zs
14の4その他:https://www.youtube.com/watch?v=AIFbGwXo1qM
15回は最終講義です。免疫系を破壊するウイルス感染症についての説明です。
15の1病原体と感染症:https://www.youtube.com/watch?v=qk2MhCuxkI8
15の2エイズ(AIDS):https://www.youtube.com/watch?v=WLPjMEUCKa4
病原体から見た感染症は、いつか「病原体の科学」で講義しましょう。
本にしたかったのですが時間がありませんでした。




生体防御系:病原体の暴露を受けた際に役立つように、宿主にはいくつかの自然バリア機構が備わっています。体表の一部を覆う粘膜には粘液(ムコ多糖+蛋白)が分泌され、病原体を凝集させたり、包んでしまうことが出来ます。また表皮は、病原体の侵入を受けにくいように、何層も重なった扁平重層上皮で覆われています。細胞内にはリゾチームがあり、細菌の細胞壁であるペプチドグリカンを加水分解して細菌を破壊します。血液中のトランスフェリン(トランス:運ぶ、フェリン:鉄、の蛋白質)は鉄と結合し、細菌が増殖するのに必要とする鉄を取り込むのを防止します。この他に体内に侵入した病原体に対抗するため、貪食能をもつマクロファージや好中球、および自然免疫(昆虫、甲殻類、原索動物まで)と特異免疫(脊椎動物の円口類、魚類以上)を発達させています。
自然免疫:その特性は、先天的に備わっている免疫系です。反応は非特異的な(病原体の持つ分子パターンを認識して反応する)生体防御反応であり、外界との接触部位(表皮、感覚器、呼吸器、消化器、泌尿器系など)に発達する傾向があります。病原体侵入時の反応は早く、炎症反応を誘起し、病原体を排除します。しかし、特異(獲得)免疫と異なり、免疫記憶は残りません。そのため、同一の病原体に再侵入されても、反応は増強しません。主なものには、生体防御レクチン(糖鎖認識蛋白)、病原体のパターン認識受容体であるトール様受容体(TLR)やNOD様受容体(NLR)、抗微生物ペプチド(ディフェンシンなど)、 C反応性蛋白(CRP、炎症時に増加し、オプソニン作用、走化性亢進作用を持ち、補体系を活性化する)、そして補体系、インターフェロン、NK細胞などがあります。

トール様受容体(TLR)とNOD様受容体(NLR): Toll-like receptor(TLR)は動物の細胞表面、細胞内にある受容体で、種々の病原体を感知して自然免疫を作動させる。TLRまたはTLR類似の遺伝子は、広く脊椎動物、昆虫、植物にも存在し、その起源は非常に古いと思われる。TLRは病原体に共通に存在し、かつ特異的な(宿主にはない)パターンを認識する。細菌表面のリポ多糖、リポ蛋白、鞭毛のフラジェリン、ウイルスの二本鎖RNA、細菌やウイルスのDNAに含まれる非メチル化CpG領域などである。Nucleotide binding oligomerization domain-like receptor(NLR)は、哺乳類では 30を超える大きなファミリーをなし、NOD1、NOD2などの5つの分子が病原微生物を認識する。NLRはTLRのように細胞表面で微生物を認識するのではなく、細胞質内で微生物を認識する。

特異免疫:自然免疫で働く血液細胞がマクロファージ(単球)や好中球、NK細胞であったのに対し、特異免疫(獲得免疫)で主体となる生体防御細胞はT,B(細胞)リンパ球である。特異免疫の特徴は、①自己と非自己(異物、病原体など)の区別をすること、②免疫記憶(病原微生物などの1回目の侵入にたいし反応すると、免疫記憶細胞が出来て、2回目以降の侵入に対し、迅速に強く反応する)が残ること、③抗原への1対1対応(抗原に対し、対応するリンパ球がクローン増殖し、抗原特異的に反応)することである。特異免疫には、免疫応答を誘導し、細胞性免疫を担うT細胞(胸腺(thymus)で分化することにちなんでT細胞と名付けられた。)と抗体産生(液性免疫応答)をするB細胞がある(鳥のファブリキウス嚢(Bursa of Fabricius)で分化するのでB細胞と命名された。)。さらに、免疫応答自体を調節する制御性T細胞があることが明らかにされた。

特異免疫の起源:これまで、脊椎動物無顎類(円口類)では、特異免疫は発達していないと考えられてきた。しかし、最近、円口類にも抗原提示細胞、T細胞に相当する細胞表面に抗原受容体を持つVLR-A細胞とB細胞の産生する抗体のようにVLR-Bを細胞外に分泌するVLR-B細胞があることが明らかにされた。ただ、魚類から哺乳類に共通するような、カセット化した遺伝子再編システムは持っていない。その意味では、魚類に至って特異免疫システムは完成したと言える(ただし魚類はIgM抗体しか産生できない。)

液性免疫:生体防御系の中で最後にできたのがB細胞を主体とする液性免疫系である。細胞性免疫系がT細胞の表面にあるTCRを介し、標的細胞に直接接触して破壊するのに対し、B細胞は抗原認識分子(BCR)を抗体として分泌し、抗原に結合させ中和反応を起こさせたり、抗原抗体反応物を補体で溶解させたり、貪食細胞に食べさせ処理させている。最も効率の良い、汎用性と特異性の高い生体防御系といえる。


インフルエンザウイルスに感染した時の生体防御反応の推移を見てみましょう。ウイルスが上部気道の上皮細胞に感染すると、レクチン系などが活性化し、炎症性サイトカインが産生され、発熱、カタール性炎、補体の活性化などが誘導されます(環形動物、ミミズのレベルくらい?)。さらにウイルスのRNAはTLRで認識されて、インターフェロンの産生が起こり、NK細胞が活性化されます(円口類、ヤツメウナギくらい)。ウイルスの増殖が続くと、下部気道の肺胞マクロファージなどが感染し、特異免疫系が活動を始めます、へルパーT細胞が細胞性免疫と液性免疫とを誘導します(魚類から哺乳類)。回復すると免疫記憶のT細胞、B細胞が残り、ワクチンと同様に、次に感染した時、速やかに2次免疫応答を起こすことができます。

第2回は、免疫現象の基盤となる特性について講義します。特異免疫応答の特徴は、①一度感染症にかかり回復すると二度がかりなし(疫病を免れるという意味で免疫という)、②自己の組織には反応しないが、他者の組織は拒絶するという、自己と非自己の認識です。③もう一つの特徴は、無限に近い種類の抗原に、1対1の対応ができる多様性がどのようにして生まれるか?です。これらの生物学的問題を解いてきたのが免疫学という学問です。

二度罹りなし:ペロポネソス戦争(紀元前431年~404年)は、アテネとスパルタの間で行われた。開戦の翌年アテネは疫病の大流行(ペスト?)に見舞われた。流行はアテネの外港ピレウスで始まり、アテネに飛び火した。不衛生な籠城生活をしていたアテネに非常な勢いで広まり、2年間で全人口の3分の1が死んだ。この時、古代アテネの歴史家ツキジデスはアテネの疫病で「2度罹りなし(疫病から回復した人は疫病に罹らない)」を記録している。同様のことは中世最大の疫病、ペスト(黒死病)でも見られ、「回復者は選ばれた人として患者の看護にあたった」とされている。麻疹についても、2度罹りなしは有名である。1846年ファロア島(Faroe Islands)で、捕鯨船乗組員から麻疹が持ち込まれ、2週間後から島内村民に麻疹が広がった。住民7800人中、およそ6100人が感染したが、老人は感染を免れた。同島の最終麻疹流行は1781年で、その時感染した住民で、今回の流行で発症する人はいなかった。すなわち、65歳以上では麻疹発症者はいなかった。






ヘルパーT(Th細胞):細胞表面にCD4抗原と抗原特異的受容体(TCR)を発現しているリンパ球の集団がTh細胞である。Th細胞は、CD4陽性T細胞から分化し、3種類のヘルパーT細胞に分化する。IFN-γ(Th1細胞)、IL-4やIL-5(Th2細胞)またはIL-17(Th17細胞)等を産生し他の細胞の活性化、機能の亢進を助ける。具体的にはTh1細胞は、主にIL-12の存在下で分化し、分化後はIFN-γを主に産生する。Th1細胞は細胞性免疫を媒介し、自己免疫疾患、遅延型アレルギーにも関与する。他方、Th2細胞はIL-13によって分化し、分化後に主にIL-4を産生する。Th2細胞は液性免疫を媒介し、即時型アレルギーにも関与する。Th17細胞は、IL17及びIL-6・TGF-β存在下で分化し、分化後はIL-17を産生する。Th17細胞は多くの自己免疫疾患モデルマウスにおいて増加していることから自己免疫疾患に関わっていることが考えられている。
キラーT(Tk)細胞:細胞傷害性T細胞(cytotoxic T lymphocyte; CTL)は宿主にとって異物になる細胞(移植細胞、ウイルス感染細胞、癌細胞など)を認識して破壊する。CTLは表面にCD8分子を発現しているT細胞から分化し、活性化されて初めて細胞傷害活性を持つようになる。細胞傷害活性を持たないナイーブCD8陽性T細胞では、T細胞受容体(TCR)が抗原提示細胞のクラスI主要組織適合抗原と共に提示された異物の抗原ペプチドを認識し、同時に共刺激分子からのシグナルが入ることで、異物の抗原ペプチドを提示する細胞に対する特異的な細胞傷害活性を持つCTLとなる。Th細胞から分泌されるIFNγなどで増殖し、標的細胞からMHCIを介して提示される抗原を持つ細胞を特異的に攻撃する。CTLが標的細胞を破壊する方法はNK細胞と同様にパーフォリンとグランザイムを利用する経路と、Fas/Fasリガンドを利用する経路があり、いずれも標的細胞にアポトーシスを誘導し、死に至らしめる。


第3回からは、人類が経験した感染症の中で、二度がかりなしという免疫現象がどのようにとらえられて来たかを、紀元前から18世紀まで振返ります。人類の脅威となったペストと天然痘の流行の影響は非常に大きいです。今回はジェンナーが二度がかりなしを種痘というワクチンの発想法で、再現性を持つ疫病の回避(天然痘ワクチン)を試みたところまでです。途中には病原体の発見につながる微生物(顕微鏡の開発により、初めて認識された生き物たち)の話も出てきます。
感染性、伝染性、伝達性の違い
感染症:より高等な動植物である宿主(host)が、病原体(pathogen) の暴露を受けた時、病原体が宿主の体内で増殖することを感染(infection)という。感染が起こった結果、疾病が生じる場合を感染症という。病原体の暴露を受けても感染しない場合もあるし、病原体が増殖しても病気にならない場合もある(不顕性感染)。
伝染病:感染症の一種。病気を起こした個体から病原体が別の個体へと伝播し、連鎖的に感染個体数が拡大する感染症を伝染病という。家畜や伴侶動物などの特定の動物種の集団内で同じ症状を示す個体が短時間に多発した状態となる。感染症法の成立後、ヒトでは使われなくなったが、家畜では使われる。例:家畜伝染病予防法
伝達性海綿状脳症:プリオン病のみに適用される。プリオン病の発病メカニズムは感染というよりも代謝異常に近い(異常プリオン蛋白の蓄積)。また、伝播様式も医原性(医療行為による伝播)、食肉習慣(牛の肉骨粉)など、通常の感染様式とは異なり人為的である。そのため感染性としないで伝達性(transmission)といわれる。伴侶動物では英国でネコの伝達性海綿状脳症が発生した。

感染症の流行に関わる3つの因子:病原体、宿主、環境
病原体:病原体は、小さい順にプリオン、ウイルス、細菌、原虫、真菌、寄生虫がある。病原体の感染力(伝播力)と病原性は、必ずしも関連しない。例えば、呼吸器感染症の場合、上部気道で増殖する病原体は伝搬しやすいが重症化することは少ない。他方、肺(下部気道)で増殖する病原体は比較的伝搬しにくいが重症化することが多い。また、病原体の伝搬のしやすさは、臓器親和性(向神経性、向呼吸器性、向泌尿器性、肝親和性など)にも関連する。増殖部位の違いは病原体の排出経路の違いとなり、感染経路の違いにも関係する(空気感染、飛沫感染、経口感染、性交感染、経皮感染、咬傷感染、母子感染(垂直感染)など)。
宿主:感染症の流行に関わる因子の1つに宿主個体の遺伝的な感受性の高さ(病原体に対する受容体の発現量の多寡)、あるいは反対に抵抗性の強さ(免疫、生体防御能の強さ)がある。また、集団で見た時には動物個体群の集団規模、飼育密度、性、年齢構成なども感染症の拡大に強い影響を与える。
環境条件:動物の飼育環境の適正化は感染症防御に重要である。特にストレスは、飼育動物の免疫機能を下げる。適当な運動、飼育温度(寒冷、温暖)、湿度、防音管理、栄養管理、衛生管理、水、換気および、病原体によっては媒介昆虫(ベクター)のコントロールも必要である。

病原体
ウイルス:ウイルスは最も単純な構造で、基本的にウイルスゲノムと複製酵素とウイルス粒子を構成する構造蛋白質からなる小さな(数十~数百ナノメートルサイズ)自己複製体である。リボゾームを持たないので、自分の遺伝情報を蛋白質に翻訳するには生きた細胞が必要となる。そのためウイルスは生きた細胞の中でしか増殖できない。
ウイルスは標的とする細胞に発現している受容体(receptor)と、ウイルス粒子表面にあるウイルス外被蛋白質(リガンド)が鍵と鍵穴のような形で合体し、細胞内にエンドサイトーシスなどで取り込まれる。あるいはウイルスの外被(脂質膜)と細胞膜が融合することで細胞内にゲノムを送り込む。ウイルスのゲノムはRNA(RNAウイルス)かDNA(DNAウイルス)であり、それぞれRNA複製酵素、DNA複製酵素でゲノムが複製される。しかし、レトロウイルス(白血病ウイルスやエイズウイルス)などはRNAゲノムを逆転写酵素でDNAに逆転写し、宿主細胞の染色体に挿入してから複製する。
ウイルスは、細菌からヒトの細胞までが2分裂で増えるのに対し、ゲノムの複製と、メッセンジャーRNAから蛋白に翻訳する過程を別々に行い、一挙に組み立てる方式をとる。そのため感染後、暗黒期(eclipse period)といわれるウイルス粒子が検出できない時期がある。組み立てられられたウイルス粒子は、細胞から出芽し、別の細胞に感染する。ウイルスにより異なるが、一般に、24時間~72時間で細胞当たり106~1011くらいのウイルス粒子が産生される。
細菌:細菌はウイルスよりも10~100倍大きな原核生物である。環状のDNAゲノム(核様体)に全ての遺伝情報が書かれている。細胞質にはリボゾーム、プラスミド、色素体、顆粒小胞、脂質顆粒などを持ち、それを細胞膜、細胞壁及び外膜(あるいは莢膜)で包んでいる。運動・移動器官などとして鞭毛、線毛を有している。細菌は細胞壁の厚いグラム陽性菌と薄い細胞壁と外膜を持つグラム陰性菌に分かれる。また、形態により、球菌(連鎖球菌、ブドウ球菌など)、桿菌、ラセン菌、その他の菌に分類される。
細菌は自分で代謝できる。光合成細菌のように自分でエネルギーを作り出す独立栄養細菌と別の生物が作った有機物を利用する従属栄養細菌がいる。この有機物を得るために毒を産生する細菌がいる。毒が産生され菌体の外に放出されるものを菌体外毒素(破傷風毒素、ボツリヌス毒素など)、菌体の内部にあって細菌が死ぬときに出る毒を菌体内毒素(リポポリサッカライド:LPSなど)という。病原性細菌は基本的に従属栄養細菌である。さらに、純培養するときに酸素を必要とする細菌を偏性好気性菌、酸素があった方がいい細菌を微好気性菌、酸素がなくても増殖できる細菌を通性嫌気性菌、酸素があると増殖できない細菌を偏性嫌気性菌という。




原虫:真核生物で動物、植物、真菌以外の単細胞生物が原生生物で、非常に多様性がある。そのうち従属栄養のものが原生動物、その中で寄生し病原性を持つものを「原虫」と定義した。細菌に比べ大型で(10μ~100μm)、ウイルス、細菌や細胞(赤血球)を貪食したり、あるいは細胞に寄生するものがある。核膜、核、核仁、ゴルジ体、ミトコンドリア、収縮胞、食胞などを持ち、鞭毛、仮足、繊毛や吸盤、口腔、食道、肛門を持つものもいる。主要な原虫には、鞭毛虫(ジアルジアなど)、根足虫(赤痢アメーバなど)、胞子虫(頭頂複合体群、トキソプラズマなど)、繊毛虫(バランチジウムなど)の4門がある。鞭毛虫と根足虫は無性生殖であるが、胞子虫と繊毛虫は有性生殖(ガメートゴニ―、小核の接合)も行う。感染経路には、経口感染、媒介昆虫(吸血昆虫)により運ばれるものが多い。
真菌:真菌は多細胞生物であり、多くは菌糸を形成するが、酵母のように単細胞のまま生活を送るものもある。主として下等真菌類に属すツボカビ門(鞭毛のある胞子形成、無性生殖)、接合菌門(接合する。菌糸の隔壁を欠く)と高等真因類(有性生殖、菌糸に隔壁を持つ)に属す子嚢菌門、担子菌門、及び不完全菌門からなる。不完全菌門は、感染し増殖しているときは単細胞性の無性生殖であり、それにより分類されたが、本来の多細胞状態でないため、菌糸形成のような多細胞状態が判明するまで不完全菌門に分類された(ヒストプラズマ、白癬菌、カンジダなど、いずれも子嚢菌)。感染病原体としての真菌はほとんどが子嚢菌である。これは担子菌門が陸生植物に寄生する傾向を示すのに対し、子嚢菌が陸生動物等に寄生して繁栄したからである。動物に病原性のある担子菌(キノコ)はほとんどいない。しかし、子嚢菌のカビ毒、キノコの中毒は起こる。
寄生虫:一般に寄生虫とは原虫から、内部寄生虫や外部寄生虫(ダニなどの昆虫を含む)まで、極めて幅広い分類である。通常は、内部寄生虫である3胚葉性の無体腔動物(扁形動物)の吸虫と条虫、偽体腔動物の線虫を蠕虫として寄生虫といっている。吸虫(肺吸虫、肝蛭など)は、吸口を持ち、多くは雌雄同体、同一個体内に卵巣、精巣、子宮を保有する(日本住血吸虫のように雌雄異体の吸虫もいる)。第一中間宿主、第二中間宿主を持つなど複雑な生活環を持つものが多い。条虫(瓜実条虫など)は、頭節と多数の体節からなり、1㎝以下から数mに及ぶものもいる。紐状の形からサナダムシともいわれ、中間宿主を持つものが多く、中間宿主内では無性生殖する(原頭節を増やす)ものがいる。線虫(糸状虫、回虫など)は脱皮する特徴を持つ。多くは雌雄異体で生殖腺を有し、交尾する。


感染症の予防
ワクチンとワクチネーション
ワクチンの多様化:19世紀の終わり頃に細菌の純培養法が確立され、また、ろ過性病原体として細菌より小さいウイルスが確認された。病原体の不活化法がわかり、生ワクチンに対して抗原性は弱いが、より安全な不活化ワクチンが作られるようになった。1920年にBCG(弱毒ウシ型結核菌ワクチン)、1930年に破傷風トキソイド、そして細胞培養法でウイルスが増殖可能となり、1955年ソークポリオワクチン(不活化ワクチン)、1957年セービンのポリオ経口弱毒生ワクチン、1960年麻疹生ワクチン、1980年組織培養型不活化狂犬病ワクチン、1983年DNA組換えB型肝炎ワクチンなどが開発された。またインフルエンザウイルスでは、不活化ワクチン、生ワクチンの他に、成分(コンポ―ネント)ワクチンも作成された。




第4回は、いよいよ19世紀後半の病原体の発見と、人工的なワクチンの開発です。ドイツにロバート・コッホ、フランスにルイ・パスツールという二人の巨人の出現により、感染症の原因と感染症の統御手段が開発されました。
感染症が病原体によって起こること、ワクチンによって発症が阻止されることが明らかになりました。しかし、何故、パスツールは、不活化ワクチンでなく、より開発の困難な弱毒生ワクチンにこだわったのか?ワクチンを作りながら、なぜ免疫現象や抗体という概念にたどり着けなかったのか?これは第5回の講義に続きます。

ワクチンの歴史
天然のワクチン:経験則から、軽い感染を起こすと本物の感染を免れるという考えは、天然痘の防御法に見られる。紀元前600年頃に中国で天然痘の痂皮を鼻腔に接種する経鼻伝痘法が確立、普及したという(人痘接種法)記録がある。種々の人痘接種法は、その後、中近東、欧米でも行われた。しかしヒト由来の天然痘ウイルスなので死亡率は1~2%と高いリスクがあった。
これに対し、ジェンナーは牛痘を用いた生ワクチン(種痘の原型)が天然痘の防御に有効であることを明らかにした。ジェンナーは乳搾りの女が「牛痘にかかると天然痘に罹らない」という言い伝えを証明しようと研究を続けた。サラ・ネルメスの手にできた牛痘の病巣(ジェンナーの調査では16症例目)を、1796年に使用人の子供ジェームス・フィップスに接種したところ、彼は天然痘の発症を免れた。これが「予防接種(種痘)」の始まりとなった。コッホやパスツールらにより、感染症が病原微生物により起こること、細菌やウイルスが、その原因であることがわかる100年近く前のことである。人工のワクチン:ジェンナーの種痘ワクチン開発に興味をもったパスツールは、自然でなく人工的にワクチンを作ることを考えた。1879年、家禽コレラ菌の培養中に、偶然、弱毒化した菌を見つけ、鶏に接種して弱毒生ワクチンとして有効であることを確認した。その後1881年に炭疽の公開ワクチン試験に成功し、次いで狂犬病のワクチン開発を進めた。パスツールは、病原体が脳や脊髄で増えていると考え、狂犬病で死んだイヌの脊髄を取り出し、ウサギの脳内に接種すると、2週間後にウサギは発症し死亡した。そのウサギの脊髄を取り出し別のウサギの脳内に接種し、これを21回繰り返した。その結果、ウサギでは発症までにかかる時間が1週間程度にまで短縮したのに対し、別の動物に対しては弱くなる傾向があることに気付いたといわれている。1885年、こうしてパスツールにより狂犬病ワクチンが開発された。ワクチン反応が免疫応答であるあることが理解されるのは20世紀に入ってからである。北里柴三郎とベーリングがジフテリア菌の抗毒素が血液中にできる抗体であることを発表したのは、1891年である。








今回は、ついに特異免疫応答の担い手が抗体であるという概念に行きつきます。
弱毒微生物と強毒微生物が互いに栄養素を取り合って干渉することで感染症を免れるという化学者の考え方から、不活化ワクチンに方向転換します。脱臭剤から消毒剤、消毒で微生物や毒素を不活化し、毒性がなくても抗原性を持つということ、そして、誘導された抗毒素(血清)が毒素を中和するという医学的発見は、ついに2度かかりなしの機構の解明に向かいました。しかし、抗体の本質を考えたのは、医学者でなく化学者のポール・エールリッヒでした。彼の抗体側鎖説はベンゼン環の修飾から生まれたといわれています。
ルーとシャンベルラン、レフレルと北里・ベーリングそしてエールリッヒと次々に天才が現れ、19世紀末から20世紀初頭に免疫学の基礎概念が成立します。






ワクチンとワクチネーション
ワクチンの多様化:19世紀の終わり頃に細菌の純培養法が確立され、また、ろ過性病原体として細菌より小さいウイルスが確認された。病原体の不活化法がわかり、生ワクチンに対して抗原性は弱いが、より安全な不活化ワクチンが作られるようになった。1920年にBCG(弱毒ウシ型結核菌ワクチン)、1930年に破傷風トキソイド、そして細胞培養法でウイルスが増殖可能となり、1955年ソークポリオワクチン(不活化ワクチン)、1957年セービンのポリオ経口弱毒生ワクチン、1960年麻疹生ワクチン、1980年組織培養型不活化狂犬病ワクチン、1983年DNA組換えB型肝炎ワクチンなどが開発された。またインフルエンザウイルスでは、不活化ワクチン、生ワクチンの他に、成分(コンポ―ネント)ワクチンも作成された。

生ワクチン:生ワクチンはウイルスや細菌などの病原体を、別の宿主で継代を繰り返し、または長期の人工培養継代などにより「弱毒化」し、生きたまま免疫に使用するものである。病原体が自然感染した時のように持続的で強い免疫が期待でき、病原性は失われているが、感染(あるいは発症)防御に必要な抗原性は保持されている。液性免疫、細胞性免疫が得られやすいが、安全で効果的な弱毒ワクチン株の作出は難しく、長期の研究が必要である。生きた抗原を投与するので、病原性復帰や変異に対する安全性の確保が必須となる。また、ワクチン株と野外の強毒株とを区別するための目印となる「マーカー」が必要である。基本的には1回のワクチネーションで強い免疫が獲得される(終生免疫に近いものもある)。
不活化ワクチン:不活化ワクチンは、ウイルスや細菌を細胞や人工培地で培養、増殖させ、増殖した病原体の液の中に含まれている培地、細胞などの不純物を除去し、熱や化学薬品で病原体を不活化したものである。不活化されているので感染は起こらず、免疫機能の低下した個体でも接種することが可能である。また、生ワクチンと異なり、母体からの移行抗体に関係なく接種可能である。しかし、細胞性免疫、液性免疫の成立が弱いので、複数回接種する必要がある(イヌの狂犬病ワクチンは、毎年のワクチネーションが義務化されている)。また、接種直後に副反応が出やすいリスクがある。

第6回は20世紀の生命科学、分子生物学をリードした免疫学の流れを中心に、100年間の進歩を1コマ(90分)で振り返るという、かなり無理のある講義です。免疫学はワクチン、抗血清療法(ベーリング、北里、エールリッヒ)から始まりますが、後の細胞性免疫の路線を敷いたウイルヒョウとメチニコフも重要です。
何故100年間の免疫学の進歩は「抗体構造から始まり、T細胞(細胞性免疫)、NK細胞、TLR、樹状細胞」のような自然免疫系へという具合に、系統発生(進化免疫)の反対方向で進んだのでしょうか?
そこには「初期発生の単純系の持つ融通無碍という複雑性、逆に、高度に複雑なシステムの持つユニットの1対1の関係という単純さ」というパラドックスがあります。分子生物学は解きやすいもとから解くという戦略であり、個別の解答を積み上げて全体を解くという帰納的アプローチです。単純系から複雑系に向かうという方法は、実際には決して楽なアプローチではありません。かなり哲学的、演繹的なアプローチになります。






NKT細胞:NK細胞とT細胞の中間に位置する、NKT細胞が見つかっている。胸腺細胞のなかで特定の受容体を選択的に発現しているT細胞が見つかり、この細胞が NK細胞マーカーを発現していることを見出した。形態学的にはNKT細胞は、NK細胞に似た顆粒リンパ球でありNK細胞とT細胞の両方の性質を合わせ持つ。



第7回からは、免疫学の各論です。進化に従って獲得した生体防御システム、すなわち自然免疫から順次獲得免疫へと展開します。第7回、8回は自然免疫系の説明です。
第7回はレクチンと補体を中心に説明します。真核細胞のうち動物細胞はαプロテオ菌に由来すると考えられるミトコンドリアを保有しています。ここで糖を利用してTCAサイクルを循環させることでエネルギー生産(ATP合成)をおこないます。動物の活動エネルギーは、基本的には、全てこの系によっています。従って、体内に糖を取り込むということは動物にとって最も基本的で必須の行為です(もっとも、糖鎖認識分子であるレクチンは、原核生物の時代から存在します。生物にとって糖は最も基本的なエネルギー源です)。
こうしたレクチンの中で、生体防御系に組込まれたレクチン群があります。パターン認識受容体の原型のような役割を演じています。また、植物のレクチンの中にはリンパ球を非特異的に活性化するものもあります。これらは免疫機能の診断にも利用されています。


原始的生体防御 ~レクチン
異物の侵入に対する防御の基本は物理・化学的なバリアーです。皮膚は細胞核をなくした扁平重層上皮からできています。積み重なった上皮細胞膜は微生物の侵入を止めます。また粘膜を覆う粘液は微生物を包んでしまうムコ多糖類や酵素などを含んでいます。リゾチームという酵素は、細菌の細胞壁を構成するペプチドグリカンを加水分解して、細菌を殺すことができます。血清蛋白のトランスフェリンは鉄と結合し、細菌が増殖のために必要とする鉄を取り込むのを防止する働きを持っています。こうした蛋白の1種にレクチンがあります。レクチンはラテン語の legereに由来する言葉で、いろいろの物の中から、「選択する、集める」という意味です。何を選別するかというと、糖を選別する、あるいは糖鎖を集めることで、現在では「糖鎖に結合活性を示す蛋白の総称」と定義されます。レクチンは細菌、真菌、植物、動物のすべてが産生しますが、高等動物ほど産生するレクチンの種類は豊富です。糖鎖はいろいろな病原体の表面などに発現しています。例えば細菌のリポ多糖体はグラム陰性菌の外膜です。ペプチドグリカンはグラム陽性菌の細胞壁の構成成分、β-グルカンは真菌の細胞壁を構成しています。ほとんどのウイルスの表面には糖蛋白が出ています。このような、病原体に共通の成分を病原体関連分子パターン(PAMPs)と呼んでいます。近年、このパターンを認識する蛋白で自然免疫に働く動物レクチンが次々と発見され、これを「生体防御レクチン」と呼ぶことが2000年に提唱されました。コレクチン,フィコリン,ガレクチン,ペントラキシンといったレクチンがこれに該当します。病原体に結合して補体を活性化したり、炎症細胞を呼び寄せたり生体防御反応の引き金として働きます。
植物レクチンには、リンパ球を非特異的に活性化するものがあります。リンパ球を非常に低濃度のレクチンと培養すると、リンパ球は突然、分裂・増殖を始めます。マイトージェン刺激と呼ばれる現象で、静止期のリンパ球を活性化する機能です。このようなレクチンにはConA(タチナタマメ)、PHA(アカインゲンマメ)、PWM(アメリカヤマゴボウ)があります。多くのマイトージェンレクチンはT細胞を活性化します。活性化したT細胞は細胞傷害性を持ち、悪性化細胞だけでなく、正常細胞も攻撃します。マイトージェンによるリンパ球活性化反応はAIDSや免疫不全の患者のT細胞機能測定に利用されます。また癌の免疫療法(LAK療法)のリンパ球分裂促進剤としても利用されています。


インターフェロン(IFN):抗ウイルス作用を持つ因子として発見された。RNAウイルスや2本鎖RNA、1本鎖RNAなどにより直接誘導される。また、種々のTLRやサイトカインによっても誘導される。IFNαとβはT細胞、B細胞、マクロファージ、線維芽細胞、血管内皮細胞、骨芽細胞など多くの細胞で産生され、特に抗ウイルス作用を示す重要な生体防御因子である。また、IFNαとβはマクロファージとNK細胞を刺激し腫瘍細胞に対して増殖抑制作用を示す。IFNγは活性化されたT細胞で産生され免疫系と炎症反応に対し調節作用を有する。特異免疫系ではTh1細胞(細胞性免疫応答系)から分泌されたIFN-γはTh2反応(液性免疫応答系)を調節する作用を持つ。
補体系:補体系は血液中の30以上の蛋白と蛋白質断片からなる生体防御系である。補体系蛋白は、通常不活性な酵素前駆体として血液中を循環しているが、どれかのトリガーで刺激を受けると、補体系の蛋白分解酵素が特定の蛋白の分解を行い、サイトカインの放出を誘導し、さらに分解反応が進むようにカスケード(前の分子が後の分子を開裂し、活性化する経路)の増幅を始める。活性カスケードの最終結果は膜侵襲複合体(MAC)の活性化による標的細胞(MACの結合した細胞)の融解を起こさせる。このように補体系の主要な役割は、抗原のオプソニン化(opsonization)、すなわち微生物などの抗原に抗体や補体が結合することにより抗原が貪食細胞に取り込まれやすくすること、膜侵襲複合体を形成し、標的とする細菌、細胞の融解を起こすこと、マクロファージ、好中球等を呼び寄せる化学遊走(chemotaxis)の3つである。また、補体を活性化する経路としては、レクチン経路、副経路、古典経路が知られている。

補体
補体は、1890年代後半に、ポール・エールリヒが抗体の側鎖説を考案し、免疫系の理論を展開した際に、生体防御系の構成要素の1つとして提案しました。新鮮な血液中に殺菌や細胞破壊など抗体の作用を補助をする因子があり、免疫反応を助けるという意味で「補体」と名づけました。1898年、パスツール研究所のジュール・ボルデは、血清中の生体防御反応が2つの要素、熱安定性要素と加熱により簡単に不活性化する易熱性要素、に分かれることを見出しました。熱安定性の要素は、特定の微生物に対して宿主に免疫を与える抗体で、易熱性要素は、感染していない正常な動物の血清の中に保持されており、 非特異的な抗微生物活性をもつことを示しました。この易熱性要素が補体でした。
補体系は単一の蛋白でなく、30種類以上に及ぶ血清中の生体防御蛋白群です。その役割は大きく3つあります。1つは、抗原のオプソニン(調理)化です。微生物などの抗原に抗体と補体が結合することにより、抗原が補体の受容体を持つ貪食細胞に取り込まれ、溶解されやすくなる現象です。2つ目は、補体蛋白の複合体(膜侵襲複合体:MAC)が細菌や細胞膜に直接穴をあけ融解させる作用です。3つ目は、マクロファージ、好中球等、炎症細胞の遊走を呼び起こす因子を産生する化学遊走(chemo-taxis)作用です。
補体系は抗体とともに発見されましたが、3つの経路が見つかりました。系統発生的には、補体系は抗体が出現する以前から存在しており、最も古いものはレクチン経路で、糖鎖を認識した生体防御レクチンが補体系を抗体なしで活性化する経路です。次いで第2経路では、細菌のリポ多糖体など種々の異物(微生物など)が、プロパージン経路を介して抗体非依存性に補体を活性化する経路です。最初に発見された古典的経路、すなわち抗体のFc部分による補体の活性化経路(抗原、抗体、補体複合体)は、皮肉にも系統発生上では最後に獲得された最も新しい経路でした。


第8回は自然免疫系の後半です、レクチンが糖鎖を、補体がレクチン系、副回路系、抗体系を介して活性化したのに対して、今回は受容体を介した初期の生体防御系である、抗微生物ぺプチチド、パターン認識によるToll様受容体(主として細胞表面の異物認識、リスク管理)、Nod様受容体(主として細胞内の異物認識、クライシス管理)とインターフェロンについて講義します。これらの受容体は外界に接する上皮細胞や、実質臓器の細胞、循環器系内皮細胞、マクロファージや好中球、あるいはリンパ球など、体内の多くの細胞に表現されています。

抗微生物ペプチド ~ディフェンシン
抗微生物ペプチド(宿主防御ペプチド)は、進化上、保存された自然免疫の1種で、生体防御機能をもつペプチドの総称です。特異免疫機能を持たない昆虫類などでは、よく発達しています。これらのペプチドは広い抗微生物スペクトルを示す一種の抗生物質で、グラム陰性細菌、グラム陽性細菌、抗酸菌、エンベロープを持つウイルス、真菌などを破壊します。
ヒトが産生する抗菌性ペプチドは大きくディフェンシン、ヒスタチン、LL37グループの3つに分類されます。ディフェンシンには、好中球が主として産生するα-ディフェンシンと上皮系の細胞が産生するβ-ディフェンシンがあります。さらに、好中球、免疫系細胞、上皮細胞は、細胞に取り込んだ細菌などを不活性化するためディフェンシンを細胞内にも持っています。ディフェンシンは、哺乳類、鳥類、魚類、節足動物だけでなく、軟体動物や線形動物にも見つかっています。抗菌ペプチドには好中球や上皮細胞などで産生されるLL37(直鎖陽イオンαへリックスペプチド)や唾液中に存在し抗真菌作用を有するヒスタチンがあります。ディフェンシンは、陽性に帯電したオリゴペプチド(18~45アミノ酸)で、弱い陰性に帯電した細菌表面などに引き寄せられ、細菌の細胞膜に貫入し、ポア(小孔)を形成し、あるいは細胞膜を破壊することで抗菌力を発揮すると考えられています。
昆虫の抗菌ペプチドとしては、細菌、真菌に幅広い抗菌作用を持つミツバチのアピデシン(18アミノ酸)、カメムシのタナチン(21アミノ酸)が知られています。また、ハチに刺されると痛いのは、メリチンというペプチドによりますが、これはカエルの抗菌ペプチドであるマガイニンやテンポリンのようにアミノ酸が26個つながったペプチドです。メリチンは細菌などが侵入したとき、細胞膜に穴をあけて細菌を壊す抗菌ペプチドです。細菌の感染防止だけではなく、細菌類を分解する効果もあるハチの重要な自然免疫機構と言えます。



パターン認識による防御 ~トール様受容体、NOD様受容体
2011年のノーベル生理学・医学賞の受賞者ブルース・ボイトラーとジュール・ホフマンは、トール(Toll)様受容体の発見者です。Toll遺伝子は1980年代にショウジョウバエで正常な発生(背腹軸の決定)に必要な遺伝子として発見されました。「Toll」はドイツ語で「偉大な」とか「素晴らしい」という意味です。1997年、哺乳類にもToll遺伝子と相同性の高い遺伝子が見つかり、トール様受容体(TLR)と命名されました。その後、TLRまたはTLR類似の生体防御遺伝子は、脊椎動物だけでなく、昆虫などにもあり、植物にも類似のものがあることがわかってきました。進化的に、その起源は非常に古いと思われます。
TLRは動物の細胞表面、細胞内にある受容体蛋白で、種々の病原体を感知して自然免疫を作動させます。獲得免疫が働くためにもTLR を介した自然免疫の作動が必要です。TLRやその他の自然免疫に関わる受容体は、病原体の進化上で保存され、常に存在する病原体に特異的なパターンを認識するものです。具体的には、TLRは細菌表面のリポ多糖体、リポ蛋白、細菌鞭毛のフラジェリン、ウイルスの二本鎖RNA、細菌やウイルスのDNAに含まれる非メチル化CpG(宿主のCpG配列はメチル化されているので区別できる)などを認識します。すなわち、TLRは特定の分子を認識するのでなく、ある一群の分子を認識するので、病原体関連分子パターン(PAMPs)認識受容体ともいわれています。
最近、TLRに似た役割を果たすNOD様受容体(NLR)が見つかりました。NLRは、哺乳類では 30を超える大きなファミリーで、そのうちNOD1、NOD2、NAIP5、IPAF、NALP3の5つが病原微生物を認識することがわかっています。NOD1,NOD2は細菌の細胞壁成分であるペプチドグリカンを特異的に認識します。しかしTLRのように細胞表面で微生物を認識するのではなく、細胞質内で微生物を認識するという特徴を持っています。硬骨魚類では数百種類のNOD様受容体スーパーファミリーが存在します。危機管理の視点からみると、TLRは主に細胞表面に病原体が来た時に活動するリスクマネージャーであるのに対し、NLRは細胞内まで病原体が来てしまったときに働くクライシスマネージャーと言えるでしょう。



抗ウイルス因子 ~インターフェロン(IFN)
IFNは、1954年、伝染病研究所(現、東大医科学研究所)所長の長野泰一と小島保彦が「ウイルス干渉因子」として発見し報告しました。1957年には、イギリスのアリック・アイザックス等もウイルス増殖を非特異的に抑制する因子としてウイルス干渉因子という意味で「インターフェロン」と命名しました。
ヒトのIFNはI型 のIFN-α(13種類)と1種類のIFN-β、IFN-ω、IFN-κがあります。II 型は1種類でIFN-γだけです。III型はIFN-λで3種類あります。IFNはTLRなどの下流にあって、病原体がTLRなどに結合すると、細胞内情報伝達系が活性化し、IFN遺伝子が開き、産生されます。IFN-α、βは分子量約2万の蛋白で、IFN-βは166個のアミノ酸からなる糖蛋白です。ヒトの成熟型IFN-γは143個のアミノ酸からできています。IFN-αとβはT細胞、B細胞、マクロファージ、線維芽細胞、血管内皮細胞、骨芽細胞など多くの細胞で産生され、特に抗ウイルス因子として重要な役割を果たします。またマクロファージとナチュラルキラー細胞(NK細胞)を刺激し、腫瘍細胞に対して増殖抑制作用も示します。IFN-γは活性化されたT細胞で産生され免疫系と炎症反応に対し誘導作用をもち、IFN-αとβの効果を増強する作用も持っています。


第9回からは、特異免疫に入ります。特異免疫(獲得免疫)の担い手はリンパ球です。特異免疫には液性免疫(B細胞)と細胞性免疫(T細胞)がありますが、特異免疫反応の基盤となる細胞は、細胞性免疫を担うT細胞(ヘルパーT細胞)です。NK(ナチュラルキラー)細胞からNKT細胞を経て、T細胞受容体を遺伝子再編により作り出す過程、調節性T細胞、免疫記憶T細胞等について説明します。



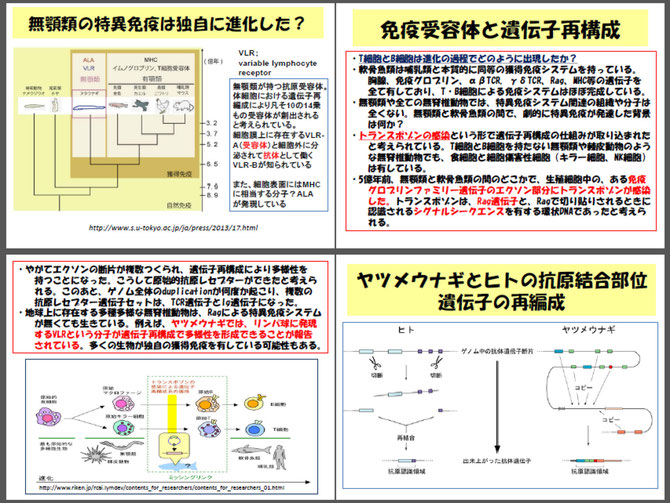

NK細胞:ナチュラルキラー細胞は、形態的には大型顆粒リンパ球(large granular lymphocyte:LGL)である。細胞内の顆粒に含まれるパーフォリンとグランザイムを使って異常になった細胞を強制的に死滅(アポトーシス)させる細胞障害機能(標的細胞を殺す活性)を持つリンパ球である。生まれつきの殺し屋細胞といえる。NK細胞が標的とするのは、主にがん細胞などの異常化した自己の細胞である。特異免疫では、クローン増殖したキラーT細胞(Tk細胞)が標的細胞の障害に同じ方法を用いる。NK細胞を活性化する因子には、ウイルス感染細胞から放出されるIFNαやIFNβ、サイトカイン(IL2)などがある。またNK細胞はFc受容体を発現しており、抗体依存性細胞傷害(ADCC)も行う。NK細胞は種々のサイトカインに応答することで、抗原特異的な細胞傷害性T細胞が生じるまでの間、ウイルス感染をコントロールするのに役立っている。

生体防御を担う細胞 ~NK細胞
生体防御を担う細胞群は、マクロファージ(大食細胞)様細胞から始まりました。生体に侵入した異物を取り込み処理する機能を持っています。系統発生的には、1胚葉側生動物の海綿動物では間細胞(原生細胞)が多分化能をもった幹細胞として機能しています。2胚葉のヒドラ、クラゲのような刺胞動物(腔腸動物)では遊走細胞は血球類似細胞ですが、貪食能はほとんどありません。同じ2胚葉性動物のサンゴ、イソギンチャクではアメーボサイトと呼ばれる遊走細胞があり、弱い貪食能を持つようになっています。3胚葉、真体腔のゴカイ、ミミズ、ヒルのような環形動物は、閉鎖血管系を形成しています。血液は血色素(巨大なヘモグロビン分子)をもち、酸素を運搬します。また、好中球、好塩基球、好酸球、顆粒球、黄細胞等の血球が血管内や体腔内に見られようになります。軟体動物などでは開放性血管系となり、血色素は銅を含むヘモシアニンです(タコ、イカは閉鎖血管系です)。血液中にはマクロファージのように外来の異物や自己由来の異物的成分を捕食し、大型異物を生体内で隔離(包囲化)する血球が存在します。凝集素や溶血素などの液性防御因子も出現します。節足動物にはリンパ球は見られませんが、ヌタウナギやヤツメウナギのような無顎類では、マクロファージ、顆粒球の他に、リンパ球が出現します。
最も古い生体防御系のリンパ球は、生まれつきの(Natural)細胞傷害機能を持つ細胞(Killer cell)という意味でNK細胞と命名されました。NK細胞が標的とするのは、主に癌細胞などの異常化した細胞です。NK細胞は、パーフォリンという細胞膜に穴をあける蛋白とグランザイムという酵素を使って異常細胞を強制的に死滅させる殺し屋細胞です。他の細胞に死をもたらす作用は、NK細胞以前のマクロファージや顆粒球にはない機能です。
NK細胞は1970年代初めに発見されました。1986年にカレらが提唱したのが自己喪失 (missing-self)説です。MHCクラスI分子は自己のマーカーで、すべての体細胞表面に発現していますが、癌細胞のようにMHCクラスI分子がない細胞であれば、それは自己性を喪失した異常な細胞であると見なし、NK細胞が攻撃するという説です。NK細胞を活性化する因子は、IFNα/β、IL-2やIFNγです。また、NK細胞は種々のサイトカインに応答することで、獲得免疫の細胞傷害性T細胞が出現するまでの間、ウイルス感染をコントロールする役割を果たします。NK細胞の発見後、15年以上たって(1986、87年)NK細胞とT細胞の中間に位置するNKT細胞が発見されました。

獲得免疫 ~細胞性免疫、T細胞
獲得免疫の特徴は、自己と非自己(異物、病原体など)の区別、免疫記憶、抗原への1対1対応(クローンリンパ球による対応)です。こうした特異性の高い免疫応答の基本的な担い手はリンパ球ですが、胸腺(thymus)で分化することにちなんでT細胞と名付けられました。
1968年、ミッチェルとミラーは、マウスの胸管リンパ液中に、抗ヒツジ赤血球IgM抗体産生細胞の前駆細胞(B細胞)と抗原依存性に抗体産生細胞に分化させる細胞(T細胞)の2つの亜集団が存在することを発見しました。その後1975年にはマラックとカプラー が T細胞の中に、免疫を誘導するヘルパーT細胞と標的細胞を傷害するキラーT細胞という機能的な違いをもつ2集団があることを報告しました。そして1986年にモスマンらが初めてマウスのTヘルパー細胞のサイトカイン分泌パターンの違いによりTh1細胞(細胞性免疫誘導)とTh2細胞(液性免疫誘導)の二つのヘルパーT細胞の亜集団の存在を報告しました。こうして、T細胞、キラーT細胞、ヘルパーT細胞、Th1, Th2というように、その機能が明らかにされてきました。
病原微生物はマクロファージ系の抗原提示細胞に取り込まれた後、抗原ペプチドにまで処理され、MHCクラスII という分子にのって細胞表面に出ます。回転ずしのお皿にのっていろいろなネタのお寿司が出てくるようなものです。トロの好きな人は、もっぱらトロ(抗原ペプチド)のみに反応します。抗原ペプチドと特異的に反応する受容体を持った(トロ好きの)ヘルパーT細胞(Th0)が反応し、クローン増殖します。その後Th1優位で免疫反応が進めばキラーT細胞クローンが増殖する細胞性免疫が、Th2優位で反応が進めば抗体産生細胞クローンが増殖する液性免疫が誘導されます。免疫反応が起こると、一部の細胞クローンは免疫記憶細胞となってリンパ組織中に残ります。ワクチンは免疫記憶細胞を誘導するために、後で本物の病原体が侵入した時に免疫記憶が活性化され、迅速に免疫反応が起こり、感染や発症を免れることができるのです。
キラーT細胞は、NK細胞と同様にパーフォリンとグランザイムを用いてウイルスや細菌などに感染した標的細胞を破壊します。NK細胞との違いは、標的細胞がMHCクラスIを使って抗原を提示し、対応する受容体を持ったキラーT細胞クローンが1対1で特異的に反応することです。パターン(PAMPs)認識による反応ではありません。


第10回は液性免疫です。分子生物学としての免疫学の花形です。抗体の構造、特異性、多様性の秘密、免疫記憶の機構などが明らかになります。また、母子免疫(移行抗体)は、胎盤構造等により、動物種差があります。
免疫現象が進化の中でどのように適応してきたかも興味ある課題です。人とマウスの免疫機構だけでなく、出来れば、それぞれの動物種の免疫機構の比較も学べるといいですね。


獲得免疫 ~液性免疫:抗体、B細胞
獲得免疫のトリの役者は抗体産生細胞(B細胞)です。基本的反応はT細胞と変わりません。T細胞受容体が細胞結合型免疫グロブリンに代わる点と、キラーT細胞が標的細胞を直接傷害するのに対し、抗体産生細胞は飛び道具である抗体を産生、分泌してウイルスが細胞受容体に結合するのを防いだり、細菌に結合し補体を活性化して溶菌させたり、様々な働きをさせます。
ほとんど無限に近い病原体の抗原に1対1に反応する抗体がどのようにできるかは免疫学の謎でした。エールリッヒの側鎖説(1つのリンパ球が多種類の抗体を作る)、ポーリングの鋳型説(抗原を鋳型として抗体が後天的に作られる)の後、バーネットがクローン選択説を提案しました。1つのリンパ球が1つの抗原に対応し、クローン増殖して1種類の抗体のみを産生するというものです。
クローン選択説は自己の抗原と反応するクローンが排除されれば、免疫系がどのようにして自己と非自己を見分けるかを説明できる魅力的な仮説です。しかし、こんなことをすれば、ヒトの遺伝子のすべては抗体分子の遺伝子で占められてしまいます。最も考えにくい仮説でしたが正解でした。謎を解いたのは利根川進で、答えはリンパ球が胸腺の中で遺伝子再編を行うという離れ業でした。個々のリンパ球は父母からもらった抗体遺伝子を再編して、一つずつ違うリンパ球になるというものです。個々のリンパ球の抗体遺伝子構造は受精卵の抗体遺伝子構造とは違っていることを証明したのです。抗体の抗原結合部位のパーツがカセットのように並んでいて、そこからランダムに取り出して組み合わせるという、順列組合せです。これだとわずかの部品で、ほぼ無限の組み合わせを作ることが可能になります。
なお、抗体は鳥類では卵黄を介して、偶蹄類では初乳を介して、ヒトやマウスでは胎盤を通過して、親から子供に移行し、新生児期の感染を防御するようになっています。



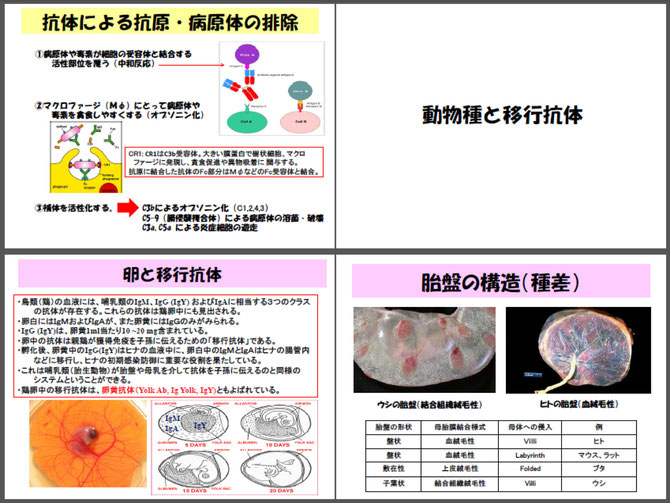
B細胞の分化:B細胞の分化は以下のように行われる。リンパ系幹細胞からプロB細胞を経てH鎖(重鎖)の遺伝子再構成が起きる。完成したH鎖とSL鎖とともにpre-BCR(プレB細胞受容体、抗体の原型)を形成、大型プレB細胞となる。pre-BCRシグナルにより一度増殖した後に、L鎖(軽鎖)の遺伝子再構成が起き小型プレB細胞へと分化する。完成したL鎖はH鎖とともにIgMを形成して、細胞膜上に発現する。IgMとともに同じ抗原特異性をもつIgDが発現する。B細胞は骨髄(鳥類ではファブリキウス嚢)から末梢へと移行し、脾臓において成熟B細胞となり、抗原の存在下で抗体を産生する形質細胞へ分化する。
遺伝子再構成:抗体分子は、H鎖とL鎖で構成され、それぞれ抗原に結合するV領域と定常のC領域からできている。V領域の多様性は、V,D,Jの3種類の遺伝子断片の積(掛け算:VxDxJ)で産み出される。免疫グロブリン(抗体)の重鎖V領域は、44個のV 遺伝子断片、27個のD遺伝子断片、6個のJ遺伝子断片から成る。軽鎖も多くのVおよびJ遺伝子断片から成るがD遺伝子断片は含まない。遺伝子再構成により免疫リンパ球は、ランダムに遺伝子断片をコピーし、約3x1011種類の抗体をつくることができる。T細胞も遺伝子再構成によりT細胞受容体(TCR)を作成する(α鎖とβ鎖)。

遺伝子再構成:抗体分子は、H鎖とL鎖で構成され、それぞれ抗原に結合するV領域と定常のC領域からできている。V領域の多様性は、V,D,Jの3種類の遺伝子断片の積(掛け算:VxDxJ)で産み出される。免疫グロブリン(抗体)の重鎖V領域は、44個のV 遺伝子断片、27個のD遺伝子断片、6個のJ遺伝子断片から成る。軽鎖も多くのVおよびJ遺伝子断片から成るがD遺伝子断片は含まない。遺伝子再構成により免疫リンパ球は、ランダムに遺伝子断片をコピーし、約3x1011種類の抗体をつくることができる。T細胞も遺伝子再構成によりT細胞受容体(TCR)を作成する(α鎖とβ鎖)。
抗体の構造と機能:抗体は、基本的にY字型の4本鎖構造(2個のH鎖とL鎖)からなり、L鎖にはλ鎖とκ鎖の2種類が、H 鎖にはγ鎖、μ鎖、α鎖、δ鎖、ε鎖の5種類がある(IgG, IgM, IgA, IgD, IgEに対応)。L鎖とH鎖はジスルフィド結合(SS結合)で結びついてヘテロダイマーを形成し、さらにこのヘテロダイマーが左右2つSS結合で結合し、Y字型のヘテロテトラマーを形成する。抗体はFab領域 (Fragment, antigen binding) 先端の部分で抗原と結合する。H鎖のFab領域とFc領域はヒンジ部でつながっている。抗原結合部位は抗原に対応するため、H鎖・L鎖ともに抗体ごとに異なっている。Fc領域 (Fragment, crystallizable)は、抗体の活性部位で、白血球やマクロファージはこのFc領域と結合できる受容体(FcR)を持っており、FcRを介して抗原と結合した抗体を認識し抗原を貪食する(オプソニン作用)。さらにFc領域は補体の活性化や抗体依存性細胞傷害作用(ADCC)など免疫反の媒介となるエフェクター機能をもっている。


第11回は特異免疫のまとめを兼ねて、免疫ネットワークの話です。ネットワークの担い手は樹状細胞(抗原提示細胞)から始まり、共通言語はサイトカイン(インターロイキン)です。クロストークする細胞はリンパ球やマクロファージです。またインターアクションする受容体分子や細胞表面分子にはCD分子や組織適合抗原(MHC分子)があります。
今回は、この辺の事情をまとめて説明します。何故組織適合抗原はH1でなくH2であったのか?スネルとゴーラの戦略の違いと、科学の真理は別の方向からせめても、一致するという感動の物語があります。ゴーラが若くして死んだためにノーベル賞をもらえなかったのは残念でした。










免疫応答はその攻撃(防疫)力のため、侵入する病原体に対しては非常に有効な武器ですが、自己認識に異常が起こると、自らを攻撃したり(自己免疫病)、また、過剰な防衛(変わった反応:アレルギー)が起こると、免疫反応自体が宿主にとって傷害となります。その意味では「両刃の剣」的存在です。第12回と13回は、自己免疫病とアレルギーについて講義します。










第13回はアレルギーです。スギ花粉症やアトピー性皮膚炎、食物アレルギーやアナフィラキシーに代表されるように、今や国民病の一つになってしまった免疫疾患です。アレ(変わった)エルギア(反応)という語源のように、異常なあるいは過敏な免疫応答です。現在では免疫反応自体が宿主に害を与える場合は、全てアレルギーに含まれます。そのため、自己免疫病もアレルギーに含まれます。アレルギーには、その発生機序によりI型からIV型まであります。代表的なものを例に説明します。










第14回は遺伝的な免疫不全症です。自然免疫系の遺伝的な不全症、特異免疫(T,B細胞)の免疫不全症などいろいろあります。その機構は、主に実験動物であるマウスで明らかにされてきました。ベージュマウス、ヌードマウス、SCIDマウス等が有名です。












第15回は感染症と免疫不全です。もっとも代表的な例は、エイズ(AIDS:
後天性免疫不全症候群)です。レトロウイルスの中のレンチウイルス感染による特異免疫不全症です。ウイルスが引き起こす免疫不全により、日和見感染症が暴発し、死に至る病です。レトロウイルスなので、一度感染するとウイルスゲノムは染色体に組込まれ、一生ウイルスを持っていくことになります。エイズウイルス以外にも、免疫系を標的にする感染症はいろいろあります。代表的な例を紹介します。期末試験は英語でやりましょう。







妻と作った人形。
娘の修学旅行の写真をもとにしました。
妻と作った人形。
娘の修学旅行の写真をもとにしました。

オリジナルの写真です

娘のドイツ時代のカーニバルの写真です。大家さんは子ネズミちゃん「モイスヒェン」といっていました。
下の人形は妻の作品です。


先日、妻の作品が創刊700号記念家庭画報大賞の佳作に入りました。
題「何して遊ぼう」です。


妻が、稽古に通い、粘土で作った作品です。昨年、東京フォーラムで、他の生徒さんと一緒に展示されました、「仙人草」
(水やり不要です)。

妻の人形作品です。
ドイツ時代の香代の幼稚園の友達です

ある夏のスナップです。妻の父母、娘、甥たちの集合写真から作りました。
